Biology
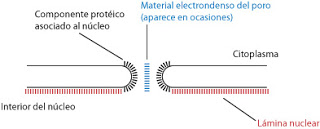
Esta membrana presenta una serie de zonas donde la membrana externa e interna se comunican, formando poros que permiten el intercambio de información entre el exterior y el interior del núcleo. Este sistema de comunicación está compuesto por el poro, es decir, el hueco y un componente protéico asociado a dicho poro.
El número de poros nucleares es variable. Son más frecuentes en los núcleos de alta actividad. Se pueden extraer complejos de poro (es decir, trozos de membrana nuclear con poros) para realizar un estudio de sus propiedades. Gracias a esto se sabe que la estructura del componente protéico asociado al núcleo es ortogonal, es decir, está formado por ocho proteínas alrededor del núcleo.
El material electrondenso d ela zona central del poro parece ser material que está atravesando el poro, acúmulos de nucleoproteínas.
Cuando se separa la zona de cabeza y de cola, la zona de la cabeza deja de pasar, mientras que la zona de la cola puede seguir pasando. Las colas se pueden asociar a oro coloidal para realizar un marcaje y vemos su capacidad de paso al núcleo.
El ARN 45s no suele verse libre, sino unido a componentes protéicos. Se visualiza como una estructura filamentosa y constituirá la zona fibrilar del nucleolo.
Parece ser que nunca se desenreda de los nucleosomas. Se piensa que el nucleosoma reconoce algún tipo de secuencia específica. La estructura de nucleosomas no es contínua, aparecen zonas enormes sin ningún nucleosoma y en otras son más frecuentes. Se cree que las zonas sin nucleosomas son oznas asociadas a proteínas de regulación génica.
- Anatomía Celular: Retículo Endoplásmico
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida...
- Formación De Arn Mensajero (arnm)
ARN polimerasas. El ADN se traduce para formar ARN, bien ARNm, ARNt o ARNr. A partir de estos tres tipos de ARN se fabrican las proteínas (como hemos visto en entradas anteriores). Las reacciones que catalizan las reacciones de fabricación del ARN...
- Estructura Básica De Los Seres Vivos.
Aparentemente, todos somos capaces de diferenciar a un ser vivo de la materia inerte. Sin embargo, esto parte de error de asociar el concepto ser vivo al concepto de movilidad, crecimiento o cambio que no siempre se corresponde con la realidad. Una rápida...
- Adn Mitocondrial: Significado Y Aplicaciones
Cuando ocurre alguna desgracia en la que toca hablar de identificación de cadáveres suele surgir un tema interesante y cuya importancia ha ido creciendo en los últimos años por diversas aplicaciones. Se trata de los análisis de ADN mitocondrial.¿Qué...
- Vih: El Virus Que Valió Un Nobel
Françoise Barre-SinoussiyLa academia sueca concedió en 2008 el premio Nobel de Medicina a Harald zur Hausen, Françoise Barré-Sinoussiy Luc Montaigner. Estos dos últimos son los investigadores que descubrieron el virus VIH (de las siglas en inglés,...
Biology
El Núcleo Eucariota.
El núcleo es una estructura englobada en los sistemas de endomembrana. Se separa de una forma muy efectiva, pero no hermética. A través de la doble membrana del núcleo existe un paso muy selectivo de sustancias.
Pudo formarse por una invaginación de la membrana de una célula procariota. En estas, habitualmente, los cromosomas están enlazadas a la membrana plasmática. También hay ribosomas enganchados a la membrana, fundamentalmente para facilitar la lectura.
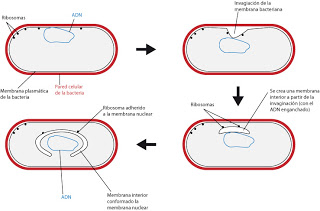 |
| Formación del núcleo eucariota en procariotas. |
La membrana nuclear no está totalmente cerrada, posee lugares de contacto directo con el citoplasma denominados poros nucleares.
Analicemos ahora las ventajas de poseer núcleo. Se relaciona con el sistema interno de ocmpartimentos y con el citoesqueleto. Los procariotas carecen de compartimentos. En los eucariotas hay sistemas de endomembrana, que facilitan que haya reacciones separadas, controlando el gasto de energía. En el caso del núcleo, este se aisla y se separan las estructuras y sustancias necesarias en los núcleo con aquellas que no hacen falta en el núcleo pero que son imprescindibles en el citoplasma. Además, en el núcleo se procesan datos. En los procariotas, cuando se traduce un gen, aun no ha acabado de traducirse cuando ya se están fabricando y formando las proteínas a partir del ARN en formación. La información extraída es la que se usa.
En el caso de los eucariotas, la información se procesa. Primero se forma el ARNm, que no se sintetiza en el núcleo. En el núcleo hay prerribosomas, que no son funcionales. Hasta que no sale al citoplasma, el ARNm no se traduce. Y hasta que sale al citoplasma, puede sufrir modificaciones. Por ejemplo, puede ser troceado en fragmentos más pequeños y que cada trozo fragmentado de lugar a una proteína diferente. Con un gen pueden ser fabricadas varias proteínas diferentes.
También hay una relación con el citoesqueleto. Debemos tener en cuenta que en procariotas no hay citoesqueleto. El citoesqueleto de los procariotas rodea al núcleo e incluso hay citoesqueleto en el interior del núcleo.
 |
| Célula eucariota con núcleo y nucleolo. |
La célula se mueve gracias a la acción del citoesqueleto. Se estira o encoge por movimientos en este cableado. El el ADN se enontrase anclado al citoesqueleto, dentro del citoplasma, los cambios de conformación prodrían romperlo, ya que es una molécula frágil. Al encontrarse en el interior del núcleo, se encuentra protegido.
La morfologíaa del núcleo es muy variada. Hay grandes variaciones en la forma, tamaño y grado de condensación de la cromatina. Tenderá a adoptar una mofología termodinámicamente estable. En muchos tipos celulares aparece con forma esferoidal. Si lo miramos con microscopía electrónica vemos que son más complicados de lo que parecen a escala fotónica. No existen núcleos de morfología realmente redondeada, presentan entrantes y salitres, en ocasiones de gran tamaño. Estos entrantes y salientes hacen que en ocasiones podamos ver orgánulos dentro de esos entrantes que, por efecto del corte, den sensación de estar dentro del núcleo (si hiciésemos cortes seriados, veríamos que está fuera del núcleo y que realmente se trata de un entrante del citoplasma en el núcleo).
Los núcleos pueden aparecer redondeados, alargados e incluso lobulados, con lo que da la sensación de existir subnúcleos (por efecto de corte, puede dar la sensación de que una célula tiene varios núcleos). Esto último es característico, por ejemplo, de los neutrófilos. El núcleo alargado es característico de las céulas musculares.
El tamaño del núcleo, como indicábamos, también es muy variable. En diferentes tipos de células hay distintos tipos de genes activos, por ejemplo, por lo cual es normal que tengan diferente tamaño y forma. En individuos de diferentes especies habrá variaciones de la cantidad de ADN. En dos células distintas de un mismo organismo, habrá diferencia de condensación. En algunas células, además, se duplica la cantidad de cromosomas (en humanos, 84 en lugar de 42) o se pierde ADN (ocurre en algunos organimos inferiores).
La variación de condensación es la causa más frecuente de la variación de tamaño del núcleo. La cromatina podrá estar formada en dos estadíos, eucromatina y heterocromatina. La eucromatina no se tiñe, mientras que la heterocromatina sí. Además, en microscopía electrónica la heterocromatina aparece con mayor electrondensidad.
Cuando se extrae información de un grupo de genes, el ADN debe estar expandido, encontrándose en forma de eucromatina. Cuando no se está extrayendo información, estará condensado gracias a la acción de ciertas proteínas (formando heterocromatina).
Esto afecta al tamaño del núcleo. Si el núcleo está en una célula muy activa, se estará extrayendo mucha información del núcleo con lo que habrá mucha heterocromatina. Esto acarrea que el ADN esté más desplegado e implicará un núcleo de mayor tamaño. Ocurre, por ejemplo, en las neuronas.
En el caso de los linfocitos, cuadno están en reposo, tienen un núcleo con el ADN muy condensado y con mucha heterocromatina. Se tiñe mucho y tiene un núcleo de un tamaño relativamente pequeño.
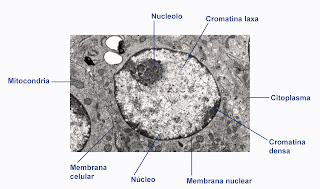
En el núcleo encontramos varias estructuras importantes. Se habla de la membrana nuclear, que lo aisla, del nucleoplasma, es decir, la parte interior del núcleo, una estructura más densa del núcleo denominado nucleolo y otras estructuras especiales denominadas esplicesoma.
Membrana nuclear.
La membrana nuclear es la encargada de aislar el contenido del núcleo. Se trata de una doble membrana, con la estructura general de una membrana biológica. Hay diferencias entre la cara externa y la interna de esta membrana. La externa es muy similar a la del retículo endoplasmático rugoso, con la que se continúa, apareciendo incluso ribosomas adheridos a su superficie (sobre todo en las zonas próximas al retículo).
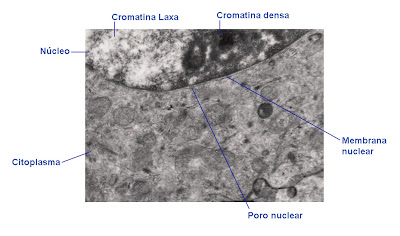 |
| Membrana nuclear con poro nuclear. |
 |
| Poro nuclear. |
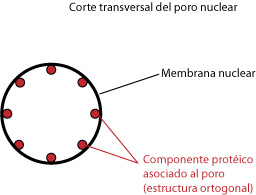 |
| Corte transversal del poro nuclear. |
Estos complejos de poro, constituidos por proteínas, están en relación con componentes asociados a la membrana nuclear interna, la lámina nuclear. Ya mencionamos algo al respecto cuando tratamos las lamininas en el citoesqueleto. No es visible en todas las preparaciones, resultando una estructura bastante difícil de observar. En ocasiones su grosor es mucho mayor que el de la membrana nuclear, en otras ocasiones apenas aparece o posee un espesor muy fino.
En la zona donde aparece un poro suele aparecer eucromatina. Da la sensación de que la zona posee una cromatina menos densa para dejar pasar las sustancias con mayor facilidad, a modo de pasillos poco electrondensos.
La unión entre las láminas y las proteínas es muy fuerte, mucho más fuerte que la unión con la membrana. Se puede lograr una extracción en la que por un lado tenemos la membrana nuclear y por otro las laminas y las proteínas del poro nuclear.
La membrana del núcleo aparece y desaparece con cierta frecuencia, ya que salvo células muy especializadas todas las células de dividen y durante la división celular la membrana nuclear debe ser degradada.
Los componentes de la láminina pueden esatr fosforilados o desfosforilados. Cuando se fosforila se desorganiza, forma dímeros y trímeros. En cambio, cuando se desfosforila se vuelven a unir para formar la lámina.
El complejo del poro supone un paso muy pequeño. Sin embargo, a través de el pueden pasar grandes proteínas. Por otro lado, el poro tiene un sistema de control del paso, siendo impermeable para muchas moléculas de pequeño tamaño. Hay una proteína en el núcleo que, cuando es extraída del núcleo y situada en el citoplasma, observamos que rápidamente regresa al núcleo. Se trata de una proteína en la que encontramos una zona de cabeza y una zona de cola.
 |
| Poro del complejo nuclear. |
Parece, por lo tanto, que las colas hacen de elemento guía, consiguen pasar al interior del núcleo cosas a las que se asocien.
También se han identificado en virus la presencia de péptids señal, muy pequeños, de unos ocho aminoácidos y ricos en lisina. Si se cambia ese péptido señal, el virus es incapaz de llegar al núcleo, se bloquea en el citoplasma.
Además, hay moléculas que nunca salen del núcleo, mientras que otras entran y salen con relativa frecuencia. Todos estos procesos están regulados. El poro hace un marcaje, una modulación del paso.
Se supone que las moléculas de gran tamaño adoptan estructuas nuevas, se alargan, cambian su conformación para conseguir pasar a través del poro.
Nucleolo.
El nucleolo puede observarse a microscopía óptica, apareciendo como una zona muy teñida dentro del núcleo. Posee morfologíaa esferoidal. Y en ocasiones no se visualiza, no son raros los núcleos que aparecen muy teñidos y sin nucleolo aparente. Pueden también aparecer núcleos con varios nucleolos, incluso con cientos de nucleolos.
Se trata de una especie de mancha denro del núcleo, con morfología variable en función del tipo celular y que carece de membrana aislante. Se divide en tres zonas, el centro fibrilar, la zona fibrilar y la zona granular. Estas zonas no siempre son distinguibles, apareciendo más claramente las diferenciadas las dos últimas y pudiendo aparecer asociadas las dos primeras.
La zona granular recibe su nombre debido a las partículas independientes que se observan en su interior. Presenta una morfologíaa variable y en general bastante extraña. Es la expresión de la actividad que está teniendo lugar en la zona del nucleolo.
El nucleolo es la zona del núcleo encargada de sintetizar el ARN de los ribosomas. Concretamente tine lugar la síntesis del ARNr, su maduración y el preensamblaje de los ribosomas.
El nuclolo es sencillamente la expresión de esta actividad. En esa zona se encuentran concentrados los genes que codifican para el ARNr. Son un grupo de genes un tanto peculiares. Para lograr una mayor amplificación en la producción del ARNr encontraremos muchas copias del gen, que suelen estar situadas secuencialmente con un espaciador intermedio. Resulta variable en número de cromosomas implicados, encontrándose especies en las que todos los genes de los ARNr se encuentran en un solo cromosoma y otras, como los huumanos, en los que se encuentran distribuidos por diez cromosomas diferentes (cinco cromosomas y sus cinco homólogos). A estas zonas de los cromosomas se les denomina organizador nucleolar y adoptan una estructura característica, acercándose entre si las zonas codificadoras y constituyendo el nucleolo.
Dada su naturaleza, es comprensible que el nucleolo pueda aperecer y desaparecer en función de las circunstancias.
Los complejos protéicos tienen como fundón fabricar ARN a partir del ADN y son fabricados secuencialmente. Primero se fabrica un ARN con un coeficiente de condensación 45s. A partir del gen se van haciendo copias sucesivas de forma continua, apareciendo una estructura en forma de árbol de navidad.
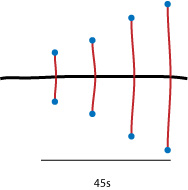 |
| Formación de ARN en nucleolo. |
Se lleva a cabo un preensamblaje de proteínas del ribosoma, fabricadas en el citoplasma, con el ARN 45s. La aparición de proteínas en la zona hace que aparezca una zona punteada, originando la zona granular del nucleolo. Hay una zona de cromatina asociada al nucleolo.
Un ribosoma es una estructura mixta, con proteínas y ARN y constituido por dos subunidades, una mayor y otra mayor, que se pueden separar. Durane el ensamblaje tiene lugar este proceso de fabricación y unión.
En el nucleolo se fabrica por un lado la subunidad mayor y por otro la subunidad menor. Según van madurando, no se permite que entren en contacto. La subunidad pequeña madura primero y sale al citoplasma (alrededor de una hora antes en la mayor parte de las céulas). En el citoplasma las dos subuniades se unen y entonces el ribosoma ya será capaz de sintetizar proteínas.
Si el ribosoma se ensamblara en el núcleo, haría que el ARN se tradujese antes de tiempo, ya que necesita ser cortado y procesado. Durante el proceso de maduración el ARN 45s es troceado, dando lugar a un ARN 28s, un ARN 18s y un ARN 5,8s. Existe también un fragmento de ARN, el ARN 5s, que se fabrica en otra zona y es indeendiente del ARN 45s.
Como decíamos, en el proceso de fabricación del ribosoma intervienen una serie de proteínas. Estas son sfabricadas en el citoplasma, fuera del núcleo y de forma independiente al nucleolo, llegando a éste, en el núcleo, a travé s de los poros nucleares. Algunas proteínas se fabrican en el citoplasma y pasan al núcleo a cumplir su función, volviendo a salir al exterior, al citoplasma, una vez han concluido su trabajo.
Otras, en cambio, son protéinas intranucleares, que aunque se fabrican en el citoplasma, pasan al interior del núcleo de donde no volverán a salir. Constituyen, por ejemplo, los bloque enzimáticos que colaboran en el procesado.
Las dos subunidades del ribosoma deben salir separadas en espacio y tiempo y se unirán, espontáneamente, en el citoplasma, formando la estructura ribosomal.
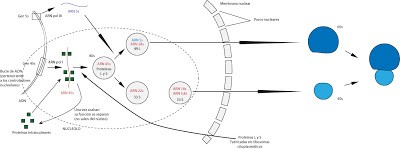 |
| Formación de ribosomas. |
Espliceosoma.
Se trata de una estructura que pued detectarse en el núcleo mediante una serie de técnicas de inmunocitoquímica, no siendo visible normalmente ni a microscopía óptica ni a microscopía electrónica. Se trata de una serie de estructuras, que apareceran marcadas mediante fluorescencia por estas técnicas inmunocitoquímicas ya citadas. Se deben a que el ARNm se mantiene un tiempo en el núcleo asociado a ribonucleoproteínas, estableciendose una serie de estructuras a modo de islas dentro del núcleo.
Nucleoplasma.
El nucleoplasma está constituido por ADN y proteínas asociadas. Cuando hablamos de proteínas asociadas, nos referimos a las proteínas que colaboran en el plegamiento del ADN para formar la cromatina o los cromosomas. Habrá otra serie de proteínas, no asociadas al ADN, que cumplirán diversas funciones en el núcleo, como complejos enzimáticos (transcriptasas, etc.).
Analizaremos la estructura de la cormatina. Es una de las partes más ingratas de la microscopía. El núcleo interfásico, altamnte activo, suele llamarse núcleo en reposo porque el ADN no está sufriendo ningún tipo de plegamiento. Solo encontramos algunas zonas más electrondensas, pero no corresponden a la realidad, la cromatina se encuentra en general asociada a proteínas y empaquetada.
Las proteínas asociadas al ADN se clasifican en dos grandes grupos, proteínas histónicas y proteínas no histónicas. Las primeras colaboran en el empaquetamiento. Las segundas son las más abundantes.
Cuando el núcleo se dsnaturaliza suavemente encontramos una hebra de un diámetro de unos 30nm. No es uniforme, cada cierto tiempo aparece una estructura redondeada, morfología que le da su nombre, estructura en collar de cuentas. Es una imagen artefactual, alterada. Normalmente el grosor sería de unos 100nm. A las estructuras redondeadas se les denomina nucleosomas. Están constituidas por ocho moléculas protéicas formadas por cuatro parejas (es decir, 4x2). Se trata de proteínas histónicas. Hay dos ploques, las histonas H1 y las histonas H2A, H2B, H3 y H4. Las cuatro parejas de los nucleosomas son las cuatro últimas (las H1 no aparecen en los nucleosomas). Se trata de una serie de proteínas muy conservadas evolutivamente (por ejemplo, las H4 de un guisante y la de una vaca solo difieren en un aminoácido).
El ADN está relacionado con el nucleosoma, de manera que da varias vueltas alrededor de este.
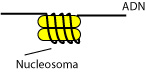 |
| Enrollamiento en nucleosoma. |
No hay datos acerca de cómo se puede formar la fibra de 100nm de grosor, que es la fibra normal en los núcleos interfásicos. Pero cuando se pasa d eun núcleo interfásico a un núcleo en división se produce un cambio, se pasa de un núcleo formado por una estructura filamentosa a un núcleo constituido por cromosomas. Los modelos que explican estos cambios son postulados teóricos.
Los nucleosomas tenderán a ponerse en contacto unos con otros, formando una estructura helicoidal, acortándose así los espacios. En esta aproximación entre los nucleosomas es donde interviene la histona H1, que no formaba parte de los nucleosomas.
En cierta medida, el modo de funcionamiento de la H1 puede asemejarse a la de la hemoglobina: cuando se le unen histonas, va adquiriendo más afinidad por el ADN.
Cuando se necesita extraer información del ADN, se retirarán los nucleosomas de esa zona en el núcleo, explicándose así que puedan leerse zonas de ADN de una forma controlada.
Para formar los cromosomas se requieren otros procesos de empaquetamiento. El siguiente empaquetamiento supone la formación de bucles. Una serie de proteínas cerrarían un lazo de ADN formando un lazo. Se trataría de proteínas de unión específica al ADN. La fibra así formada tendría un grosor de unos 300nm, todavía muy lejos del grosor de un cromosoma.
En el último grado de empqueamiento los conjuntos de fibras se ascian entre si formando otro bucle más amplio. Y este grosor si se aproximaría al grosor del cromosoma.
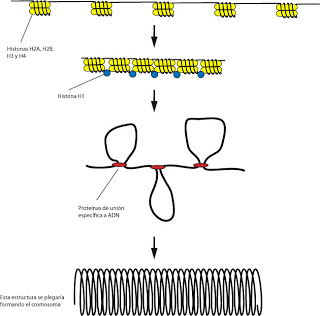 |
| Enrollamiento para formar cromosomas. |
- Anatomía Celular: Retículo Endoplásmico
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida...
- Formación De Arn Mensajero (arnm)
ARN polimerasas. El ADN se traduce para formar ARN, bien ARNm, ARNt o ARNr. A partir de estos tres tipos de ARN se fabrican las proteínas (como hemos visto en entradas anteriores). Las reacciones que catalizan las reacciones de fabricación del ARN...
- Estructura Básica De Los Seres Vivos.
Aparentemente, todos somos capaces de diferenciar a un ser vivo de la materia inerte. Sin embargo, esto parte de error de asociar el concepto ser vivo al concepto de movilidad, crecimiento o cambio que no siempre se corresponde con la realidad. Una rápida...
- Adn Mitocondrial: Significado Y Aplicaciones
Cuando ocurre alguna desgracia en la que toca hablar de identificación de cadáveres suele surgir un tema interesante y cuya importancia ha ido creciendo en los últimos años por diversas aplicaciones. Se trata de los análisis de ADN mitocondrial.¿Qué...
- Vih: El Virus Que Valió Un Nobel
Françoise Barre-SinoussiyLa academia sueca concedió en 2008 el premio Nobel de Medicina a Harald zur Hausen, Françoise Barré-Sinoussiy Luc Montaigner. Estos dos últimos son los investigadores que descubrieron el virus VIH (de las siglas en inglés,...