Biology
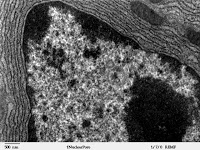 Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida celular, si lo encontraremos en alguna de sus fases. Hay células que se diferencian en un momento dado y que pueden llegar a perder todo o parte de su retículo, aunque en su fase juvenil si que aparecerá en relativa abundancia; esto ocurre por ejemplo en los eritrocitos, o los espermatozoides (en sus fases adultas presentan muy poco retículo, ya que en su maduración lo pierden, ya que en sus fases juveniles sí que presentan retículo en abundancia).
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida celular, si lo encontraremos en alguna de sus fases. Hay células que se diferencian en un momento dado y que pueden llegar a perder todo o parte de su retículo, aunque en su fase juvenil si que aparecerá en relativa abundancia; esto ocurre por ejemplo en los eritrocitos, o los espermatozoides (en sus fases adultas presentan muy poco retículo, ya que en su maduración lo pierden, ya que en sus fases juveniles sí que presentan retículo en abundancia).
Las diferencias entre el retículo liso y el rugoso se encuentra en que el rugoso presenta una serie de estructuras, a modo de gránulos electrondensos, pegados a la membrana. Estos gránulos son en realidad otro orgánulo celular asociado: ribosomas. Además, la morfologíaa general del retículo rugoso es un poco diferente al del liso, ya que el rugoso está formado por cisternas aplanadas, alargadas, comunicadas entre si por conexiones más estrechas (pudiendo aparecer también ciesternas independientes). Los ribosomas siempre están anclados a la hemimembrana que mira hacia el citoplasma, es decir, hacia el exterior del retículo (al encontrar zonas con muchas cisternas paralelas, puede inducir a error).
En el retículo liso no hay ribosomas. Además, se trata de estructuras tubulares, forma una especie de madeja de tubos.
Componentes del retículo endoplasmático.
El ribosoma detiene su trabajo hasta que llega el retículo. Sin embargo, hay excepciones. En algunos organismos este proceso no se lleva a cabo de esta forma, sino que se realiza la síntesis en el citoplasma y posteriormente la proteína es dirigida al retículo.
Los péptidos señal también se usan para indicar qué trozos de la proteína van a quedar en contacto con la membrana. Puede tener uno o varios trozos hidrófobos. Cuando aparece un trozo hidrófobo al final del la proteína, como se indica en el esquema, se está señalizando que la proteína quedará anclada en la membrana. Si esto ocurre, el extremo amino de la proteína quedará siempre hacia el interior de la membrana del retículo y el extremo caboxilo quedará hacia la parte exterior.
Pero en ocasiones las proteínas están en la membrana en el otro sentido, con el grupo amino haia el exterior y el carboxilo hacia el interior. Esto se consigue incluyendo un péptido señal hidrófobo a mitad de la proteína, quedando este incluido a mitad de traducción y forzando a salir al exterior al extremo amino.
Las protéinas transmembranales con varios pasos y múltiples hélices se integran gracias a la aparición de múltiples secuencias de péptido señal. Debemos tener en cuenta que las secuencias de péptido señal irán apareciendo duante el proceso de síntesis, es decir, durante la propia fabricación de la proteína en el retículo.
En el retículo se forman la mayoría de las proteínas de membrana, tanto de la plasmática como de otros orgánulos como el Golgi.
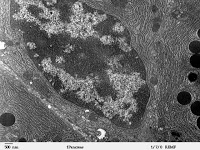 Esta biosíntesis lipídica se da en todas las células vivas. Las membranas deben movilizarse y deben renovarse. La fabricación de hormonas, en cambio, solo se da en algunos tipos celulares, como las células del testículo o la corteza suprarrenal. Esta biosíntesis suele vanir asociada con mitocondrias de crestas tubulares, resultando un símbolo inequívoco de que la célula fabrica hormonas y lípidos esteroideos.
Esta biosíntesis lipídica se da en todas las células vivas. Las membranas deben movilizarse y deben renovarse. La fabricación de hormonas, en cambio, solo se da en algunos tipos celulares, como las células del testículo o la corteza suprarrenal. Esta biosíntesis suele vanir asociada con mitocondrias de crestas tubulares, resultando un símbolo inequívoco de que la célula fabrica hormonas y lípidos esteroideos.
- El Núcleo Eucariota.
El núcleo es una estructura englobada en los sistemas de endomembrana. Se separa de una forma muy efectiva, pero no hermética. A través de la doble membrana del núcleo existe un paso muy selectivo de sustancias. Pudo formarse por una invaginación...
- Aparato De Golgi Y Lisosomas
Características del Aparato de Golgi. Camilo GolgiEl aparato de Golgi es un tipo de orgánulo visible solo bajo el microscopio electrónico. Está constituido por membranas. Fue identificado de forma indirecta mediante estudios de microscopía...
- Membranas Celulares.
Hay muchos tipos de membranas celulares. Son algo más que sistemas de la célula:Forman o aíslan compartimentos cerrados. Regulan el paso o movimientos de sustancias a través de ellas. Se encargan del paso de información: las membranas...
- Breve Aproximación A La Fisiología Muscular.
Los músculos son los encargados de producir los movimientos del cuerpo. Además de facilitar los movimientos de unas partes del cuerpo respecto a otras, los músculos permiten otro tipo de movimientos, como los movimientos involuntarios del intestino...
- Vih: El Virus Que Valió Un Nobel
Françoise Barre-SinoussiyLa academia sueca concedió en 2008 el premio Nobel de Medicina a Harald zur Hausen, Françoise Barré-Sinoussiy Luc Montaigner. Estos dos últimos son los investigadores que descubrieron el virus VIH (de las siglas en inglés,...
Biology
Anatomía Celular: Retículo Endoplásmico
Se trata de una estructura que se suele dividir en dos grandes tipos o grupos, el retículo liso y el rugoso. No hay una diferencia o falta de continuidad neta entre ambos, se trata de la misma estructura aunque con una estructura, composición lipídica y protéica que les confiere unas ciertas características diferenciales. Pero realmente se trata de una estructura común, que si pudiésemos extender comprobaríamos que uno y otro se encuentran conectados. Además, en algunas células encontraremos conformaciones de retículo particulares, que no se adaptan a ninguno de estos dos tipos.
 |
| Tipos de retículo endoplásmico |
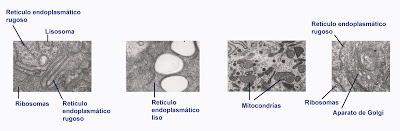 |
| Orgánulos celulares |
Función del Retículo Endoplasmático.
retículo endoplasmático puede aislarse con relativa facilidad. Para su estudio se fragmentan las células, induciendo que se separen las membranas interiores. El retículo tenderá a cerrarse sobre si mismo, formando microsomas. Estos microsomas son activos y de esta manera ha podido analizarse su función.
Para esta serie de estudios se realiza la extracción recurriendo a células con mucho retículo. Normalmente se eligen hepatocitos (células del hígado), que poseen gran cantidad de retículo tanto liso como rugoso. El retículo rugoso es especialmente fácil de separar, ya que presenta mucha densidad debido a los ribosomas. En un gradiente de densidad, se separa formando una banda bien definida. En el caso del retículo liso, no se separa con tanta facilidad y por eso debe recurrirse a una célula con mucha cantidad de retículo liso, de forma que sea más probable su aislamiento.
El retículo interviene en la biosíntesis de proteínas, encargándose de fabricar proteínas transmembranales de los orgánulos celulares y de la membrana celular. También fabrica proteínas de los lisosomas. Esta función es desarrollada básicamente por el retículo rugoso.
También interviene en la biosíntesis lipídica. Son exportadas, posteriormente, a los sistemas de membranas celulares. De esto se encargará, fundamentalmente, el retículo liso.
Por otro lado hay una función de glicosidación. Se encargaría de una glicosidación parcial de las proteínas. Se para en un estadío de glicosidación concreto y después en el aparato de Golgi se completará el trabajo. Es decir, en el retículo endoplasmático, concretamente en el rugoso, comienza la glicosidación de las proteínas y los lípidos.
También posee una cierta función de detoxificación. Esta función corre a cargo del liso. De ahí que se encuentre muy desarrollado en células epaticas. El retículo realiza una transformación de ciertas moléculas para que puedan ser metabolizadas o eliminadas por el sistema del limpieza del organismo. Por ejemplo, en el caso de sustancias hidrófobas, capaces de fusionarse con la membrana, puede ser transformada para convertirla en una molécula hidrófila, que pasará al citoplasma donde podrá ser eliminada o limpiada. A veces esto puede resultar un arma de doble filo, hay sustancias que no son especialmente dañinas y al ser modificadas por el retículo se transforman en sustancias tóxicas o cancerígenas.
El retículo es una estructura que puede crecer y decrecer rápidamente. Un tóxico, como los barbitúricos, que son eliminados por el retículo, provocan la proliferación del orgánulo. Este hecho se ha aprovechado para aislar retículo, ya que si suministramos barbitúricos a un animal de laboratorio, nos aseguramos de que el retículo sea más abundante en los hepatocitos. Cuando quitamos los barbitúricos de la dieta del animal, el retículo se reduce rápidamente, siendo digerido o reciclado por la propia célula.
El retículo endoplasmático liso también interviene en procesos metabólicos como la glucogenolisis, es decir, la degradación del glucógeno. Esta función es especialmente evidente en los hepatocitos (el hígado es una de las reservas de glucógeno).
Finalmente, interviene en procesos de secuestro de calcio. El calcio es una molécula importante que interviene en muchos fenómenos, desde la adhesión celular hasta la contracción muscular. En las células musculares, por ejemplo, hay acúmulos de un retículo especial denominado retículo sarcoplásmico (a las células musculares se les denomina sarcómeros). Se trata de una modificación del retículo endoplasmático liso, que posee en su membrana una ATPasa asociada encargada del transporte de iones de calcio. Cuando se necesita una contracción, este calcio que se acumula en el retículo se libera al citoplasma. La concentración de calcio intracelular es menor que la extracelular, el calcio está muy controlado. Hay, en todas las membranas de los componentes celulares y en las membranas del retículo transportadores de calcio que controlan su concentración. No se sabe si existen especializaciones del retículo liso en todas las células o si esta función, en células normales, es llevada a cabo por un retículo endoplásmico liso normal. La proteína identificada encargada de acumular el calcio se denomina calciosecuestrina, que es capaz de inmovilizar el calcio dentro de las cisternas.
Componentes del retículo endoplasmático.
La membrana del retículo es ligeramente más estrecha que la plasmática. Su composición es normal, pero existe una gran diferencia en cuanto a las proteínas de membrana que encontramos en la membrana celular respecto a la del retículo. Hay proteínas relacionadas con el anclaje de los ribosomas, denominadas genéricamente riboforinas (no está claro que se trate de un solo tipo de proteína). Provocan una unión mediada por fuerzas débiles (aunque al anclaje también colabora el péptido en formación cuando el ribosoma del retículo está trabajando).
En el retículo endoplásmico no hay continuidad, hay una diferenica protéica. No hay movimientos de proteínas desde el retículo rugoso al liso. Puede ser que se formen microislas de proteínas asociadas, lo cual hace que la movilidad se reduja. O bien se forman anclajes a otras proteínas. No obstante, se trata solo de hipótesis, sin que se sepa realmente qué mecanismos son los que actúan.
Hay proteínas que se forman en el interior del retículo y alguna parte de la proteína formada tendrá algún tipo de señal que hace que no pueda salir del interior. Otras proteínas se fabricarán fuera y entrarán o difundirán al interior. Y un grupo de proteínas se asociará a la membrana del retíclo. El marcaje es necesario para explicar que algunas proteínas nunca se muevan del retícuo.
En la membrana del retículo liso también hay protéinas, destacando dos citocromos, el B5 y el P450. Son cadenas de transporte de electrones, parecidos a las de la mitocondria. Pero no se produce respiración, ni fosforilación oxidativa. Se trata de un sistema para metoxilar compuestos, realizando por ejemplo reacciones relacionadas con la detoxificación.
Biosíntesis protéica asociada al Retículo.
Hay dos grandes ciclos de síntesis protéica, a nivel citoplasmático o en relación con las membranas del retículo endoplasmático. Existe otro ciclo menor y paticular de síntesis protéica, asociado a algunos orgánulos especiales, principalmente en las mitocondrias y cloroplastos.
La biosíntesis de proteínas en el retículo conlleva una serie de procesos previos que indicarán a la maquinaria si la proteína tiene que quedarse en la membrana del retículo. En cualquier caso, comienza en el hialoplasma. Los ribosomas capturan el ARNm en el hialoplasma. Lo consiguen gracias a lo que se denomina péptido señal. Cuando una proteína se fabrica en el laboratorio por medio de ribosomas libres, se obtiene una proteína de mayor longitud de lo normal. Debe existir, por lo tanto, algún trozo del péptido que se perdía entre la tradución y la síntesis en los ribosomas. Lo primero que sintetiza es la señal de reconocimiento. En el hialoplasma hay partículas de reconocimiento que se unen al péptido señal y al ribosoma. Bloquean la zona e inducen el desplazamiento del conjunto al retículo endoplásmico rugoso. En el retículo hay un apartícula que reconoce a la partícula de reconocimiento del hialoplasma. De este modo, se une al conjunto. La primera partícula de reconocimiento se desprende y vuelve al hialoplasma.
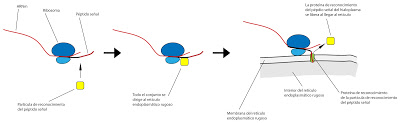 |
| Fabricación de proteínas en el retículo |
El sistema del péptido señal permite identificar algunos tipos de proteínas. Puede darse el caso de que existan varias eñales. Si tenemos una protéinas, en forma de cadena polipeptídica, la señal de unión a la membrana del retículo endoplásmico estará constituida por aminoácidos hidrófobos. Esa zona, ese trozo, marcará la zona transmembranal de la proteína. De forma que una proteína puede tener una ovarias de estas zonas de señalización, de forma que podrá atravesarl la membrana una o varias veces. Existen varias posibilidades. El péptido puede soltarse de su señal, quedando por lo tanto encerrada en el interior del retículo. O no soltarse de la señal, quedando por lo tanto la proteína enganchada o atravesando la membrana del retículo. En el siguiente esquema se omite el ribosoma, para simplificar el esquema.
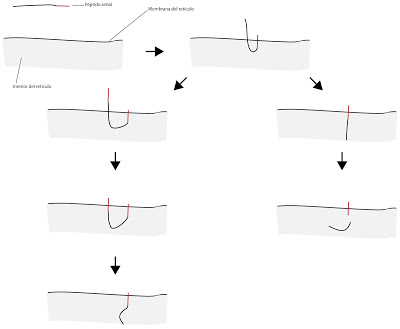 |
| Sistema del péptido señal |
Pero en ocasiones las proteínas están en la membrana en el otro sentido, con el grupo amino haia el exterior y el carboxilo hacia el interior. Esto se consigue incluyendo un péptido señal hidrófobo a mitad de la proteína, quedando este incluido a mitad de traducción y forzando a salir al exterior al extremo amino.
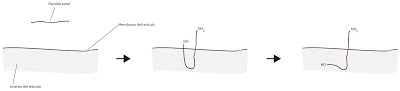 |
| Péptido señal: extremo amino en el exterior |
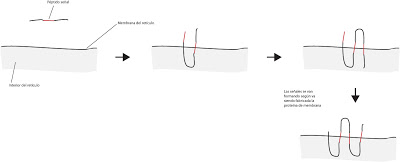 |
| Péptido señal en proteínas transmembranales |
Se consideraba que el péptido señar era muy específico, pero hoy se considera que no lo es tanto. Sencillamente, se juega con zonas hidrofóbicas. Comienzan a fabricarse péptidos con péptidos señal, que al ser zonas hidrofóbicas se van insertando en la membrana.
El plegado de proteínas y la unión de varias proteínas entre si es un proceso problemático. La proteína, al ser fabricada de forma lineal y ser una molécula con radicales cargados, debería unirse a otras proteína o moléculas cargadas y precipitar. Se conocen dos sistemas para evitar estas combinaciones indeseadas entre proteínas. Una de ellas se encuentra en la cavidad del retículo y separa unas proteínas de otro, sobre todo aquellas que tienden a unirse. Otro sistema que se ha identificado se encargaría de controlar la formación de puentes disulfuro. Existe una proteína en el retículo que se encarga de romper los puentes disulfuro inadecuados, facilitando o ayudando a que se formen proteínas con los puentes disulfuro adecuados.
La glicosidación también es un fenómeno muy importante. Las características de una proteína depende de su glicosidación. Se realiza en el Golgi, pero comienza en el retículo. Se hace un marcaje de las proteínas que deben ser glicosidadas. Se ponen una serie de 14 azúcares, que actuará como árbol de azúcares identificativos. El árbol de azúcares se va uniendo según se va formando la proteína.
En el retículo no hay azúcares libres. Hay una ruta general, se unen a una base mediante gasto energético. Así formamos el azúcar activado. El dolicol, que se encuentra en la membrana del retículo endoplasmático rugoso, es el aceptor del azucar. El dolicol hace un movimiento flip-flop introduciendo el árbol de azúcares en la cavidad del retículo. Se rompe el enlace del árbol de azúcares con el dolicol y así queda introducido en el interior. Es un proceso difícil de entender, sobre todo a nivel energético.
Otro proceso que tiene lugar en el interior del retículo es la fabricación de proteínas que se unen a fosfolípidos mediante enlaces no muy fuertes. Se fabrican, por ejemplo, las proteínas encargadas de unirse al fosfatidil inositol. La función de estas proteínas no es muy conocida. Puede ser que se unan para destruir marcadores. Se sueltan con relativa facilidad. Un ejemplo clásico son la N-cam, que median en fenómenos de adhesión celular en las células nerviosas. Harían desaparecer proteínas de la superficie celular.
En el retículo también tienen lugar procesos de biosíntesis lipídica. Tiene lugar en el retículo endoplasmático liso. Hay dos grandes bloques de rutas, las que foman los lípidos de membrana y las que fabrican hormonas esteroideas y se encargan de la biosíntesis del colesterol.
Con respecto al resto de lípidos, su fabricación se lleva a cabo hacia el citosol. La biosíntesis solo se produce en la hemicapa que mira al citoplasma. Esto acarrea una desigualdad de membrana importante, cada cara de la membrana debe tener unas características especiales. Para lograr esta diferencia hay procesos de flip-flop de lípidos. Cambian de cara en la bicapa Lippidica. Existen unas proteínas que se encargan de transportar a los lipidos de un lado a otro de la bicapa, aunque no son ni bien conocidas ni se han aislado en muchos casos. Es el sistema que logrará equiparar los lípidos.
Los lípidos son necesarios para todos los sistemas de membrana. Todos estos sistemas están en contacto con el aparato de Golgi y con el retículo. Las mitocondrias no entran dentro de este sistema, ya que son orgánulos muy característicos, con su propio material genético incorporado y ribosomas para fabricar buena parte de sus proteínas. La mitocondria no tiene biosíntesis lipídica, a la mitocondria no llegan vesículas, posee proteínas especiales encargadas de recoger ciertas proteínas del retículos, que las protegen (las proteínas hidrófobas que necesita la mitocondria no pueden vagar por el citoplasma, debido precisamente a su carácter) y las llevan a la mitocondria.
Las membranas del retículo endoplasmático se continúan con la membrana nuclear. La membrana externa del núcleo es una continuación del retículo en la que, en muchas ocasiones, podemos observar ribosomas adheridos (debemos recordar que el núcleo es una doble membrana, con una parte externa y una interna).
- El Núcleo Eucariota.
El núcleo es una estructura englobada en los sistemas de endomembrana. Se separa de una forma muy efectiva, pero no hermética. A través de la doble membrana del núcleo existe un paso muy selectivo de sustancias. Pudo formarse por una invaginación...
- Aparato De Golgi Y Lisosomas
Características del Aparato de Golgi. Camilo GolgiEl aparato de Golgi es un tipo de orgánulo visible solo bajo el microscopio electrónico. Está constituido por membranas. Fue identificado de forma indirecta mediante estudios de microscopía...
- Membranas Celulares.
Hay muchos tipos de membranas celulares. Son algo más que sistemas de la célula:Forman o aíslan compartimentos cerrados. Regulan el paso o movimientos de sustancias a través de ellas. Se encargan del paso de información: las membranas...
- Breve Aproximación A La Fisiología Muscular.
Los músculos son los encargados de producir los movimientos del cuerpo. Además de facilitar los movimientos de unas partes del cuerpo respecto a otras, los músculos permiten otro tipo de movimientos, como los movimientos involuntarios del intestino...
- Vih: El Virus Que Valió Un Nobel
Françoise Barre-SinoussiyLa academia sueca concedió en 2008 el premio Nobel de Medicina a Harald zur Hausen, Françoise Barré-Sinoussiy Luc Montaigner. Estos dos últimos son los investigadores que descubrieron el virus VIH (de las siglas en inglés,...