Biology
 Todas las células vegetales presentan una pared celular rígida, además d euna familia de orgánulos propios, denominados plastos o plastidios (entre los que encontramos los cloroplastos). Por otro lado, tiene otro orgánulo específico, llamado vacuola vegetal y que resulta una especie de almacén celular. Carecen de centiolos ni estructura derivadas, como cilios o flagelos.
Todas las células vegetales presentan una pared celular rígida, además d euna familia de orgánulos propios, denominados plastos o plastidios (entre los que encontramos los cloroplastos). Por otro lado, tiene otro orgánulo específico, llamado vacuola vegetal y que resulta una especie de almacén celular. Carecen de centiolos ni estructura derivadas, como cilios o flagelos.
Los componentes más antiguos se presentan en la zona periférica. Los más recientes están en estrecha relación con la membrana plasmática.
En la pared secundaria hay tres estratos diferenciados. Del exterior al interior se denominan S1, S2 y S3.
Las pectinas so polímeros de hidratos de carbono, ramificados y localizados en los espacios intermedios entre las fibras de celulosa. Se unen a las porciones libres de la hemicelulosa. De esta manera, las microfibrillas de celulosa se unen gracias a la hemicelulosa.
Las ligninas son moléculas derivadas del fenol. Se trata de polímeros formados por la unión de alcoholes fenólicos, principalmente alcohol coumarílico, coniferílico y sinepílico.
Se trata de compuestos químicos muy hidrófobos. La lignina es exclusiva de paredes secundarias. Impermeabiliza la pared. Esto supone matar a las células y por eso la lignina es propia de células en las que ya no hay citoplasma ni núcleo.
Génesis de la pared.
Estas vesículas, que constituyen lo que se denomina fragmentoplasto, comienzan a unirse entre si. Se van fusionando y en este proceso, se va formando la membrana intermedia. Lo que está en el interior de las vesículas formará el componente del espacio intermembrana.
Una gran parte de las características de la célula vegetal vendrán determinadas por la pared celular. En algunos estudios solo se analiza la pared celular. Para aislarla introducimos el tejido en hipoclorito y así se elimina el citoplasma.
El desmotúbulo puede variar su diámetro. Cuanto mayor sea el desmotúbulo, mayor control del paso de sustancias, ya que por los lados dejará menos espacio de citoplasma para el paso libre de sustancias. A través del desmotúbulo pasan multitud de sustancias. Pueden pasar incluso partículas virales. Es un paso mucho más ancho que, por ejemplo, las uniones Gap.
En las punteaduras aeroladas hay una cavidad, la pared secundaria se eleva y se produce, de esta forma, un redoble. Como indicábamos, estas pueden ser también unilaterales o dobles. El espacio que queda bajo la elevación se le denomina cavidad de la punteadura y al canal, poro de la punteadura.
Debemos tener en cuenta que estos dibujos son esquemáticos, pues realmente la pared secundaria es mucho más gruesa que la primera, que actuaría como membrana de cierre.
Los plasmodesmos se originan durante el primer estadío de formación de la pared. En determinados puntos, la fusión de las vesículas tras la mitosis deja huecos, pudiendo observarse que determinados elementos del retículo endoplasmatico se van quedando en el medio, formando la esturctura que será el desmotúbulo.
Los cloroplastos presentan una estructura características, con una membrana plastidial externa y una membrana interna. En los cloroplastos la membrana interna no tiene invaginaciones (como ocurre con las mitocondrias), son lisas. Dentro hay unas membranas que forman una especie de láminas aplanadas, acumulándose en forma de sacos alargados. A cada estructura se le llma tilacoide. Al conjunto de tilacoides se denomina granos o grana.

Una vez se ha obtenido esa energía, la primera reacción consiste en que una molécula de dióxido de carbono se une a una molécula de ribosa (ribulosa 1, 5 difosfato).
La ribulosa 1,5 bifosfato carboxilasa cataliza la primera reacción y se considera el enzima más abundante de la Tierra. El proceso es lento, con unas tres moléculas por segundo. Por eso debe haber muchas moléculas de enzima. Supone alrededor del 50% del contenido del estroma del cloroplasto. Está constituido por varias cadenas polipeptídicas. La mayor de ellas está codificada por el ADN propio del cloroplasto.
Los ribosomas del colorplastos presentan índices de sedimentación 60S. Se asemejan mucho al de los procariotas (una prueba más del origen simbionte de estos orgánulos). Y el ADN se transmite solo desde la parte femenina de la planta (como ocurre con las mitocondrias).
- Tejidos Vegetales: Epidermis.
Características generales. Cubierta vegetal: epidermis y tricomas. (por Louisa Howard)Es un tejido de revestimiento, siempre en la superficie de todos los tejidos. Es el tejido de revestimiento primario, aparece solo en las zonas donde hay un revestimiento...
- Histología Vegetal: Meristemos.
Características.Tejido Vegetal (Foto: Doc. RNDr. Josef Reischig, CSc.) Se trata de un tejido constituido por células pluripotenciales, indiferenciadas. Atendiendo a su localización topográfica, tenemos meristemos apicales, cuando se encuentran...
- Fotosíntesis
Introducción. La energía de la luz acaba transformándose en la energía de los compuestos orgánicos. Estos serán utilizados como hemos visto en temas anteriores, bien directamente o indirectamente mediante cadenas trópicas. La energía de la luz...
- Anatomía Celular: Retículo Endoplásmico
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida...
- Breve Aproximación A La Anatomía Celular.
La célula es la unidad funcional de los seres vivos; excepto algunas excepciones de seres muy simples Célula Eucariotacomo virus y priones, los organismos vivos están compuestos por una o más células. Cada célula es, en si mismo, un ser vivo capaz...
Biology
La célula vegetal.
Introducción.
Estas son las principales características que diferencian las células animales de la vegetales.
Pared celular.
En los tejidos no todo el espacio está ocupado por células. Hay un medio externo. En la parte inferior de muchos tejidos tanto animales como vegetales existe una membrana, compuesta fundamentalmente por proteínas y que se denomina lámina basal.
La pared celular de vegetales es algo similar, pero de mayor tamaño y la rodea completamente. Su grosor puede variar desde 0,1?m hasta 10 ó 20?m. No es deformable y hace que las células vegetales sean totalmente inmóviles. No vamos a tener ni células similares a las musculares ni tejidos en general especializados en el movimiento.
La pared celular presenta una estructura con tres estratos. El primero se denomina lámina media, la segunda se denomina pared primaria y la tercera la pared secundaria.
La lámina media es compartida por las dos células y constituye su sistema de adhesión. La pared primaria se presenta cuando la célula está creciendo, aumentando su volumen. La pared secundaria es más gruesa y cuando aparece, normalmente, la célula perderá su contenido citoplasmático y su núcleo.
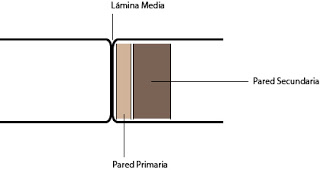 |
| Estructura de la pared celular |
En la pared secundaria hay tres estratos diferenciados. Del exterior al interior se denominan S1, S2 y S3.
Cada estrato posee una composición química propia. En la pared primaria el componente más habitual es un polisacárido, la celulosa. Son cadenas de glucosa unidas por enlaces ?1-4 (otros polisacáridos, como el almidón, son cadenas de glucosas unidas por enlaces ?1-4 y en estos enlaces radica la diferencia que hace que no podamos digerir madera, mientras que los rumiantes sí pueden, pues estos poseen en su estómago protozoos con enzimas capaces de romper los enlaces de la celulosa). Se trata de una cadena lineal sin ramificaciones. Se establecen puentes de hidrógeno entre cadenas paralelas. Con esto se consiguen haces muy gruesos y resistentes, siguiendo una determinada pauta.
La pauta es la siguiente: 32 cadenas de celulosa se asocian por puentes dehidrógeno y se va formando lo que se llama una fibrilla elemental. 20 fibrillas elementales se asocian para dar lugar a una microfibrilla. 200 microfibrillas se asocian para dar lugar a una fibrilla. Y 2500 fibrillas se asocian y dan lugar a una fibra de celulosa.
En la pared primaria toda la celulosa está en forma de microfibrilla. En la pared secundaria estará en forma de fibrillas o de fibras de celulosa.
La celulosa no es el único componente de las paredes. Otro componente típico son las hemicelulosas. Son polímeros de celulosa. En ellas encontramos uniones ?1-4, pero son moléculas muy cortas y con ramificaciones. Se asocian a las fibras de celulosa.
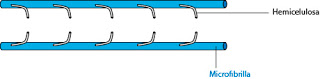 |
| Celulosa y hemicelulosa |
Las glucoproteínas aparecen a concentraciones más bajas. Son proteínas ricas en aminoácidos de serina, treonina e hidroxiprolina, es decir, con grupos hidroxilo libres.
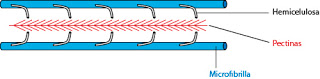 |
| Celulosa, Hemicelulosa y Pectinas |
 |
| Ligninas |
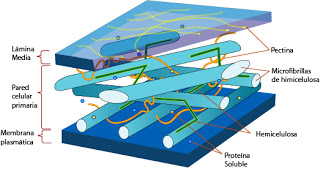 |
| Estructura tridimensional de la pared primaria |
Génesis de la pared.
Las hemicelulosas y pectinas se sintetizan en el aparato de Golgi. Estos compuestos no requieren de una secuencia específica de azúcares; en cambio, las glucoproteínas sí requieren de una secuencia específica. Esto significa que para su generación sólo se necesita un tipo de enzimas: los enzimas que catalizan la formación de cadenas de glucosa con enlaces ?1-4. Son proteínas integrales de membrana. Van al aparato de Golgi y de ahí salen en vesículas hacia la zona donde estará la lámina media.
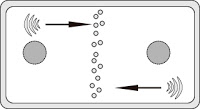 |
| Génesis de la pared |
Las vesículas, que vienen del aparato de Golgi, tienen pectinas y hemicelulosas. Van a formar una pared muy rica en pectinas, constituyendo lo que será la lámina media entre ambas células. Su función es mantener las células unidas, pegadas.
La membrana plasmática tiene una serie de proteínas de membrana con los componentes necesarios para formar la celulosa. Las vesículas provenientes del Golgi comienzan después a aportar materiales necesarios para fabricar la celulosa.
Del aparato de Golgi provienen vesículas con los materiales necesarios. Hay un vertido de cadenas lineales de celulosa. Las cadenas de celulosa se van asociando en el exterior. En un primer momento, las cadenas pequeñas, constituidas por microfibrillas, se asocian de una forma bastante azarosa. Dan lugar a una estructura anisótropa, formando una especie de pared formada por agujas sin orientación alguna. Esta estructura constituye la pared primaria.
Si la célula es de una zona que requiere actividad citoplasmática, como una hoja, esta será la única pared que aparecerá. Pero si la función del tejido es, por ejemplo, mantener erguida la planta, las células perderán su citoplasma en su estado más adulto.
Este proceso se debe a la acumulación en la pared de un aporte de material de forma continua. La celulosa comienza a estar constituida por muchas menos cadenas de celulosa, pero mucho más largas. Se formarán fibras de celulosa. Estas no se pueden entrelazar si se disponen al azar y por eso comenzarán a tomar una estructura espacial preferente. Se orientan de una forma mucho más ordenada.
La pared secundaria presentará por este motivo un aspecto fibroso. También presentará entre sus componentes pectinas y otros componentes. Finalmente comenzará el aporte de lignina. Esta no proviene del aparato de Golgi, sino de lisosomas y de algunas derivaciones de las vacuolas. La lignina impermeabilizará la estructura.
La pared celular funcionará ahora como una barrera de permeabilidad. Esto es algo secundario, las moléculas de gran tamaño no la pueden atravesar, las de pequeño tamaño sí. No obstante, se trata de una pared muy rígida. Actuará como un exoesqueleto, es decir, como un esqueleto externo.
Las concentraciones salinas interiores de la célula son mucho mayores que las exteriores en las células vegetales. El agua tiene tendencia a entrar en grandes cantidades. La célula no aumenta de volumen y explota gracias a la pared. Si quitásemos la pared, la célula aumentaría de volumen hasta explotar.
Por este motivo la membrana plasmática está pegada a la pared, ejerciendo sobre ella una presión. Se denomina presión de turgencia o turgar. Esa presión será la responsable de guiar el crecimiento y los posibles movimientos de las células vegetales.
El turgor controla el tamaño y la forma de la célula. Si la pared fuese homogénea, la célula tendería a adquirir una forma esférica. En determinadas zonas, en cambio, hay partes más rígidas, mientras que otras partes son menos rígidas. Esto marcará las zonas de crecimiento de la célula vegetal.
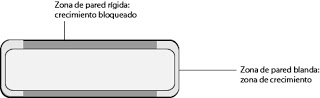 |
| Zonas de crecimiento de la pared |
La presión de turgencia es muy importante. En algunas células puede llegar a las 50 atmósferas.
Incorporación de nuevos materiales.
Se han descrito cuatro posibles mecanismos para incorporar nuevos materiales:
- Aposición.
- Intususpección.
- En mosáico.
- Multirreticular.
La aposición propone que los materiales se superpondrían a los ya existentes, sin unirse a los anteriores y formando capas nuevas, que empujan a las otras a partes exteriores.
La intususpección consiste en que los materiales no quedan alineados o formando una capa, sino que se acoplan parcialmente a la capa ya formada. Los materiales nuevos y ya formados se unen y reorganizan.
El mecanismo en mosáico propone que en determinadas ocasiones hay una desordenación parcial de la pared y se podrían, en determinadas regiones, generar una ruptura. Es en estos huecos donde se añaden nuevos materiales.
En el mecanismo multirreticular los materiales ya existentes tienen capacidad de reorganizarse. Debido a la presión de turgencia, se produce reorganización, los componentes se colocan de forma más paralela. La organización ocuparía un volumen menor, los materiales nuevos no están orientados por lo que ocupan más volumen. Debido a la presión de turgencia estos se volvería a organizar y ocupar menos volumen dejando huco para nuevos materiales.
No está claro qué mecanismo se cumple, en algunas células parece que uno y en otras parece que otro. Los dos primeros son los principales, los dos últimos no dejan de ser variaciones de los dos primeros.
Las paredes de las células vegetales presentan discontinuidades, interrupciones. Se denominan punteaduras. Hay punteaduras primarias o primordiales en la pared primaria y sencundarias o punteaduras sin más en la pared secundaria.
Las punteaduras primarias son mucho más pequeñas y se descubrieron más tarde. Son muy importantes y están relacionadas con los plasmodesmos. Las células con solo pared primaria solo tendrán punteaduras primarias.
Las punteaduras son huecos y no deben confundirse con los plasmodesmos. En los plasmodesmos existe una continuidad de las dos membranas, los citoplasmas están unidos. El plasmodesmo está ocupado por una estructura en forma de i mayúscula, que forma lo que se denomina desmotúbulo. El desmotúbulo se continúa con el retículo endoplásmico rugoso.
La punteadura supone una continuidad física entre las células y supone un paso sin control de sustancias a ambos lados de las células. En cambio, en el plasmodesmo hay un control de paso a través del desmotúbulo, se trata de una unión más específica, solo pasan algunas moléculas.
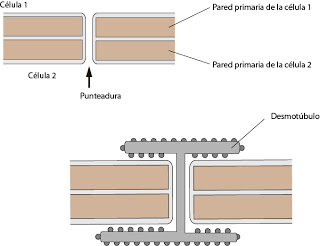 |
| Punteaduras y plasmodesmos |
Todas las células con protoplasto tienen este tipo de estrucuturas, en algunos tipos celulares pueden ser especialmente abundantes. Todas las cavidades internas de las células vegetales están, por lo tanto, unidas.
Debido a esto se habla de simplasto, cuando nos referimos a la cavidad formada por todas las células. Un producto químico puede pasar de una célula a otra muy alejada sin salirse del citoplasma.
Nos referimos al apoplasto cuando hablamos de la cavidad formada por el exterior de todas las células vegetales, sin tner en cuenta el interior.
Hay zonas con muchas punteaduras primarias y se habla de campos de punteaduras.
Las punteaduras secundarias o punteaduras propiamente dichas, suelen darse entre células sin protoplasto. En este caso aperecerá una membrana de la punteadura o membrana de cierre, que es la zona de pared primaria y lámina media que está en la zona de la punteadura. Al espacio abierto en la pared secundaria se le denoimina cavidad de la punteadura. El hueco en sí mismo es la punteadura secundaria. Las punteaduras secundarias pueden ser simples o aeroladas. Y dentro de cada uno, podemos encontrar punteaduras dobles (normales) o unilaterales.
Comencemos con la estructura de una punteadura simple:
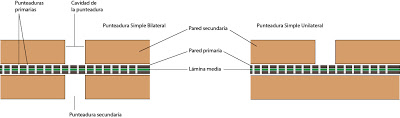 |
| Punteadura simple |
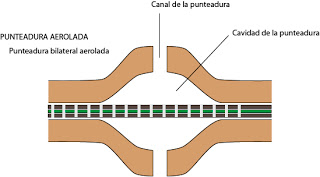 |
| Punteadura aerolada |
Es habitual que en estas punteaduras aeroladas exista un engrosamiento de ma membrana de cierre. Se le denomina Toro. Funciona a modo de válvula, sube y baja ante diferencias de presión. Se trata de una región impermeable. Al deformarse, subir y bajar tapa el poro, obliterando el orificio o canal de la punteadura. De este modo se controla el paso del agua de célula a célula.
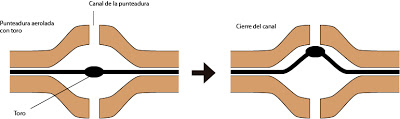 |
| Punteadura aerolada con toro |
Plastidios.
En las células vegetales hay un orgánulo denominado plastidio o plasto. Se trata realmente de un conjunto o familia de orgánulos con doble membrana. Poseen genomia propio, ADN, e incluso ribosomas propios específicos, resultando de esta forma que actúan como orgánulos semiautónomos. Todos provienen de un mismo origen, de un protoplasmidio.
Dentro del concepto de plastidio hay varios tipos, pudiendo hablar de cloroplastos, cromoplastos y leucoplastos.
Los cloroplastos tienen pigmentos tipo clorofila y en ellos se realiza la fotosíntesis.
Los cromoplastos son plastos parecidos a los cloroplastos, pero con otros pigmentos. Usan longitudes de onda diferentes.
Los leucoplastos no tienen color, no poseen pigmentos. Tienen funciones de reserva, es decir, de almacenamiento. Pueden almacenar almidón y hablamos de amiloplastos, o proteínas y hablamos de proteoplastos. Cuando acumulan lípidos se les denomina oleoplastos.
Que el protoplastidio vaya a formar uno u otro tipo de plasto depende de los primeros estadíos de formación, de los primeros pasos. Hay una cierta posibilidad de interconversión.
Por ejemplo, as patatas poseen gran cantidad de amiloplastos. Al ponerlos al sol, transformarse en cloroplastos y la patata adquirirá coloraciones verdosas.
Los plastidios son orgánulos con una cierta independencia, pues por ejemplo son capaces de dividirse de manera autónoma.
Cloroplastos.
 |
| Cloroplasto |
El DNA es una molécula circular, pequeña. Presenta un número alto de copias de, entre 5 y 15 aunque este número varía bastante. Suele presentar una zona de unión con la membrana interna.
La clorofila y sus proteínas se encuentran en la membrana de los tilacoides, donde fomran una especie de granos.
Es en esta zona donde se realizará el proceso de fotosíntesis. La reacción global de la fotosíntesis se puede resumir de la siguiente forma:
Se pueden distinguir dos etapas, una en la que la célula obtiene enrgía gracias a la luz, acumulándola en forma de ATP y NADPH, y una segunda en la que, a partir de ese ATP y NADPH, da lugar mediante una serie de reacciones , a la formación de componentes ricos en energía (glucosa, azúcares, etc.).
La primera reacción de la fotosíntesis ocurre en las membranas tilacoidales. El segundo paso se incia en el estroma de los cloroplastos y se completa en el citoplasma de la célula.
Encontramos dos fotosistemas, el I y el II. El fotosistema II obtiene la energía de la luz a 680nm, mientras que el fotosistema I obtiene energía de la luz a 700nm. En la caída de energía de los electrones se bombean al espacio intermembrana, como en la mitocondria. El flujo de electrones se usa para obtener moléculas energéticas.
 |
| Fotosistemas |
 |
| Reacción de la ribulosa 1, 5 difosfato carboxilasa |
 |
| Cloroplasto inactivo |
El funcionamiento es diferente cuando hay y cuando no hay luz. Cuando no hay luz, el cloroplasto no solo no funciona, sino que además varía su morfología. Adquieren una morfología característica y se les denomina etioplastos. También presentan alteraciones bioquímicas, bajando los niveles de algunos enzimas como el 1,5 BP Carboxilasa. Se trata de un proceso reversible, recuperando los pigmentos y niveles enzimáticos cuando la luz aumenta.
La vacuola vegetal.
Es un orgánuylo muy espcífico, que no se parece en nada a las vacuolas animales. Almacenan productos ricos en energía, agua, sales minerales, etc. Pueden ocupar mucho volumen dentro de la célula. De hecho, en algunos casos contribuyen a que el volumen de la célula sea grande. Esto es importante en células fotosintéticas, ya que a mayor volumen, mayor exposición a la luz.
Presentan una membrana denominada tonoplasto. Se trata de una membrana con muchas proteínas de transporte específicas. Se cree que deriva del aparato de Golgi.
El proceso de formación de las vacuolas recuerda al proceso de formación de los lisosomas. Del aparato de Golgi salen vesículas marcadas que llegan a la membrana de la vacuola. La membran de la vesícula se incorpora de esta manera a la membrana vacuolar.
Los carbohidratos que acumulan suelen ser polímeros, como el almidón. Es importante en el proceso de regulación de la presión de turgencia. El almidón, en estado polimérico, es una molécula osmóticamente inactiva. Pero la glucosa sí que es activa. La despolimerización del almidón es importante para controlar esta presión osmótica, por lo tanto. Al despolimerizarse el almidón, aumentamos la presión de turgencia.
La vacuola no está relacionada con la expulsión de materiales al exterior, sino para la acumulación de materiales en el interior.
Ausencia de centrilos.
Las células de vegetales claras, evolucionadas, es decir, a partir de algas. La ausencia de centriolos implica que las células, normalmente, no son móviles, les falta un sistema para remover el medio. También implica diferencias en cuanto al citoesqueleto. En las células animales, el aparato de Golgi está formado por sáculos que se separan unos de otros y se mantienen en esa posición gracias al citoesqueleto, por ejemplo (de este modo, si añadimos una sustancia que desorganice los microtúbulos, el aparato de Golgi desaparece). En las vegetales hay moléculas de unión intercisternar, es decir, poseen un sistema de anclaje diferente.
En la división celular no hay un estrangulamiento, sino que se forma un fragmentoplasto, a partir de vesículas fabricadas en el aparato de Golgi. Por lo que no hace falta organización de citoesqueleto.
Metabolismo secundario.
Las células vegetales tienen metabolismo secundario. Consiste en reacciones de síntesis de productos que, en muchas ocasiones, no se conoce su función, como alcaloides, etc. (por ejemplo, el caucho). En ocasiones son productos de desecho, en otras feromonas.
Aparecen en el citoplasma en forma de inclusiones de diversa naturaleza. Las células animales, en ocasiones, también tienen inclusiones, aunque no son tan frecuentes ni abundantes.
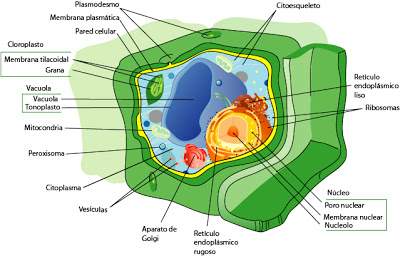 |
| Esquema de una célula vegetal |
- Tejidos Vegetales: Epidermis.
Características generales. Cubierta vegetal: epidermis y tricomas. (por Louisa Howard)Es un tejido de revestimiento, siempre en la superficie de todos los tejidos. Es el tejido de revestimiento primario, aparece solo en las zonas donde hay un revestimiento...
- Histología Vegetal: Meristemos.
Características.Tejido Vegetal (Foto: Doc. RNDr. Josef Reischig, CSc.) Se trata de un tejido constituido por células pluripotenciales, indiferenciadas. Atendiendo a su localización topográfica, tenemos meristemos apicales, cuando se encuentran...
- Fotosíntesis
Introducción. La energía de la luz acaba transformándose en la energía de los compuestos orgánicos. Estos serán utilizados como hemos visto en temas anteriores, bien directamente o indirectamente mediante cadenas trópicas. La energía de la luz...
- Anatomía Celular: Retículo Endoplásmico
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida...
- Breve Aproximación A La Anatomía Celular.
La célula es la unidad funcional de los seres vivos; excepto algunas excepciones de seres muy simples Célula Eucariotacomo virus y priones, los organismos vivos están compuestos por una o más células. Cada célula es, en si mismo, un ser vivo capaz...