Biology

- Anatomía Celular: Retículo Endoplásmico
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida...
- Metabolismo: Cadena Respiratoria.
En entradas anteriores hemos analizado las fases iniciales del metabolismo de la glucosa: la glucolisis y la fosforilación oxidativa, junto con los mecanismos de degradación anaerobia y el Ciclo de Krebs. En este paso analizaremos la cadena respiratoria....
- Transporte A Través De Membrana
Transporte por difusión física simple. Es un transporte pasivo y no mediado. La membrana debe ser permeable para el producto en cuestión, que pasará desde la zona más concentrada a la zona más diluida. En teoría, cualquier molécula se puede disolver...
- Adn Mitocondrial: Significado Y Aplicaciones
Cuando ocurre alguna desgracia en la que toca hablar de identificación de cadáveres suele surgir un tema interesante y cuya importancia ha ido creciendo en los últimos años por diversas aplicaciones. Se trata de los análisis de ADN mitocondrial.¿Qué...
- Vih: El Virus Que Valió Un Nobel
Françoise Barre-SinoussiyLa academia sueca concedió en 2008 el premio Nobel de Medicina a Harald zur Hausen, Françoise Barré-Sinoussiy Luc Montaigner. Estos dos últimos son los investigadores que descubrieron el virus VIH (de las siglas en inglés,...
Biology
Fecundación animal.
Fecundación y gametos.
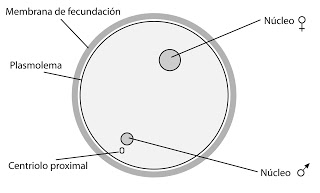
Los gametos tienen poco tiempo de vida, solo unas horas. El espermatozoide debe llegar al óvulo lo más rápido que pueda y cederle los n cromosomas de su núcleo. Además, aportará al cigoto un centriolo (es decir, los centriolos proceden del padre). El espermatozoide hace que el óvulo en parada metabólica inicie su desarrollo.
Veremos como ejemplo de fecundación el caso del erizo de mar. El macho vierte los espermatozoides en el agua de mar. Deben alcanzar al óvulo. Además, el espermatozoide no debe fecundar a otro tipo de célula. Es decir, necesitamos que sea específico.
El encuentro tiene lugar al azar, pero el movimiento de los espermatozoides siempre termina por encontrar algún óvulo. En algunas especies tiene lugar una atracción química.
Primer reconocimiento específico.
El primer reconocimiento específico tiene lugar cuando el espermatozoide entra en contacto con la ganga. Hay una reacción de interacción entre las glucoproteínas de la ganga y las de la membrana en la zona del acrosoma. No es una interacción totalmente específica. Puede ocurrir que existan reconocimientos incluso de otras especies de erizo de mar.
 |
| Primer reconocimiento específico |
Se va a producir la activación o capacitación del espermatozoide, ara que pueda entrar dentro del óvulo. Es desencadenado por el reconocimiento. Y se produce lo que se denomina reacción acrosomal.
Reacción acrosomal.
Tiene dos componentes la exocitosis del acrosoma y la formación del tubo acrosómico.
En la exocitosis del acrosoma se produce una reacción en la membrana del espermatozoide, permitiendo la entrada de iones de calcio por unos canales abiertos por ligando. El calcio induce, por un lado, la fusión de las membranas del acrosoma y del óvulo. Como consecuencia de esta fusión de membranas, se libera el contenido del acrosoma.
 |
| Liberación del material del acrosoma |
Entre las sustancias liberadas del acrosoma están las lisinas espermáticas, que hidrolizan la ganga.
Al mismo tiempo, el calcio activará en la membrana una bomba de intercambio que introduce sodio (Na+) en la célula y expulsa protones (H+), es decir, transporte antiporte. El sodio entra por gradiente de concentración, ya que hay mucho sodio en el agua de mar (recordamos que hablamos de la fecundación en erizos de mar). La expulsión de protones hace que el pH del interior suba. Al subir el pH se produce la polimerización de filamentos de actina, que hace que se forme un tubo denominado tubo acrosómico. En la membrana que recubre el tubo acrosómio hay una proteína, que inicialmente miraba hacia adentro del acrosoma y ahora mira hacia fuera y se denomina bindina.
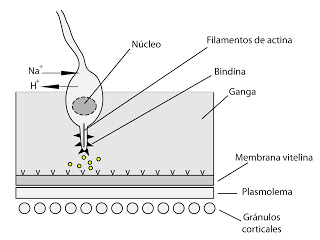 |
| Formación del tubo acrosómico |
En resumen:
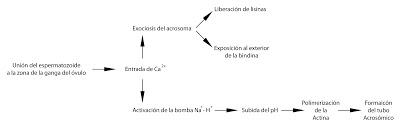 |
| Resumen de la fase temprana en la fecundación. |
El tubo acrosómico aumenta de tamaño y avanza ayudado por la lisina. Toca la membrana del óvulo. Entonces se producen un reconocimiento específico entre la bindina y unos componentes receptores de la membrana del óvulo. Estamos en la fase de reconocimeinto específico.
Reconocimiento específico.
Hasta aquí podrían haber llegado varios espermatozoides a la vez a un mismo óvulo. Pero a partir de aquí, a partir del reconocimiento específico, solo puede avanzar uno.
El sistema que veremos es parecido al que tiene lugar en mamíferos, aunque no es exactamente igual. Debemos tener en cuenta que en los óvulos de mamíferos no hay ganga, hay otra serie de líquidos diferentes. Habría exocitosis acrosomal, pero no se formaría tubo acrosómico y la fusión, en lugar de ser por un tubo acrosómico (que no tiene) se lleva a cabo lateralmente, directamente con la membrana del espermatozoide. Pero también habrá, lógicamente, un reconocimiento específico.
Como decíamos, en el erizo de mar el tubo llega a la membrana del óvulo y la bindina se une a los receptores.
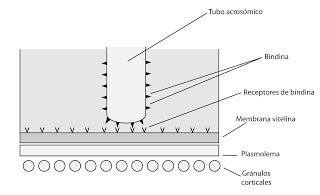 |
| Reconocimiento específico del proteínas de bindina |
Tras el reconocimiento, tendrá lugar la fusión de membranas.
Fusión de membranas.
Se unen la membrana del espermatozoide con el plasmolema.
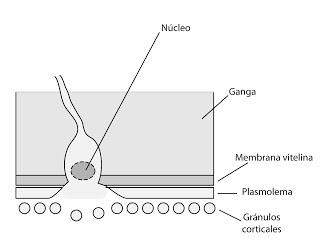 |
| Fusión de membranas |
El óvulo, tras esto, forma un pequeño abultamiento en forma de verruga en esta zona, que se denomina cono de fecundación. Y comenzará a penetrar en el interior, es decir, habremos llegado a la fase de penetración.
Penetración.
El cono de penetración empuja al espermatozoide hacia adentro. Pero el espermatozoide no entra al completo, lo que queda dentro es el núcleo y el centriolo, mientras que el flagelo permanece en el exterior. El cono de penetración está formado por una especie de microvellosidades.
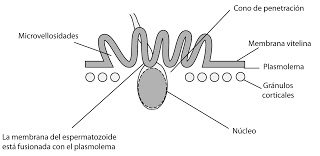 |
| Penetración del núcleo del espermatozoide en el óvulo |
Solo un espermatozoide puede entrar dentro del núcleo (monoesperma), solo se puede fusionar la membrana de un espermatozoide. Puede llegar más de un espermatozoide a la fase de reconocimiento, pero no entrará más que un espermatozoide. En caso contrario, se formaría poliespermia, con lo cual el resultado sería una célula 3n. Lo normal sería que esta célula no sería viable, moriría. No obstante, debe haber un sistema que evite la poliespermia. Y para ello, el óvulo debe ser activado.
Activación del óvulo.
El óvulo se encontraba parado metabolicamente. Y el espermatozoide lo saca de este estado. Desencadena el programa de desarrollo del óvulo, que se encuentra impreso en el propio óvulo. Experimentalmente, se puede lograr que el óvulo se desarrolle sin ser fecundado, por ejemplo pinchando con una aguja la membrana. Al desarrollo natural de los óvulos sin ser fecundados se denomina partenogénesis y en algunas especies es un mecanismo normal (en ocasiones, hay gametos que son 2n).
La activación del óvulo comienza tras el reconocimiento específico. Este sería un punto sin retorno. Y encontraremos dos grandes tipos de respuestas al reconocimiento, las respuestas tempranas, que suceden en el primer minuto tras el reconocimiento, y las respuestas tardías, que suceden después del primer minuto.
La primera respuesta temprana es un cambio en la concentración de sodio. En menos de un segundo se producen la apertura en la membrana del óvulo de un canal de Na+. Dado que es un ión abundante en el agua de mar, está mucho más concentrado que en el interior del óvulo, con lo que se produce la penetración del mismo.
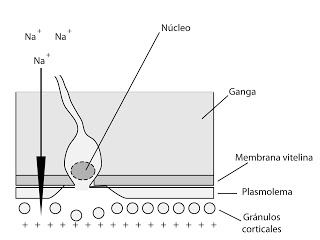 |
| Intercambio iónico en la fusión de membranas |
La membrana del óvulo posee, originalmente, un potencial transmembrana negativo. Tras la apertura de los canales de sodio, este entra en el interior a favor de gradiente y provoca un cambio en el potencial transmembrana, llegándose a valores de 20mV. Es decir, la membrana sufre una fuerte despolarización. Si la membrana no está polarizada, no se puede fijar ningún espermatozoide más, debido probablemente a algún cambio en la membrana. Y si había algún otro espermatozoide enganchado (pero sin fusionar su membrana) ahora ya no podrá entrar, ya no podrá fusionar su membrana.
Lo más probable es que en cuanto se fusione uno, ya no se puedan fusionar más. Pero hay un cierto porcentaje de errores en los que penetran más espermatozoides de la cuenta. Si antes de que se fusione el primer espermatozoide se producen una despolarización artificialmente, ningún espermatozoide podrá entrar.
En unos minutos, la membrana volverá a su estado normal, así que debe establecerse otro bloqueo para que ningún espermatozoide entre más tarde. Y si denominamos al anterior primer bloqueo, ahora comenzaremos a hablar de segundo bloqueo de la poliespermia.
Se formará la membrana de fecundación. Sucede entre veinte y treinta segundos más tarde. Hay un aumento del calcio (Ca2+) dentro del citoplasma. Procede del propio óvulo. Se encuentra dentro de los gránulos corticales. Los gránulos corticales liberan calcio al interior de la célula y este aumento de calcio induce a los gránulos a fusionarse con la membrana del óvulo y verter su contenido al exterior por exocitosis. Al liberar calcio y fusionarse con la membrana, los granos inducen a los granos vecinos a llevar a cabo el mismo proceso, es decir, a liberar calcio al interior y liberar su contenido al exterior por exocitosis. Y esto induce también la liberación en los vecino. Y de este modo la señal se va propagando por todo el óvulo.
 |
| Liberación de los gránulos corticales |
El óvulo pasa de amarillo a blanco. Debemos tener en cuenta que el óvulo tiene, aproximadamente, 15000 gránulos corticales.
Entre los componentes que se liberan al exterior hay proteínas y glucopolisacáridos. Un enzima va cortando las uniones de la capa vitelina con el plasmolema, con lo que ambas se van separando. Y otro enzima destruye la glicoproteína de los receptores de bindina.
Los glucopolisacáridos hacen que entre agua, por procesos osmóticos, a este espacio que se genera entre la membrana vitelina y el plasmolema. Se va generando un espacio perivitelino. Una serie de proteína estructurales se incorporarán a la membrana vitelina. Esto provoca que se haga más gruesa, pasando a denominarse membrana de fecundación. La membrana de fecundación establece el segundo bloqueo.
Otra serie de glucopolisacáridos se quedarán unidos al plasmolema, generando la capa hialina. Esto ayuda a que durante las primeras divisiones las células queden pegadas unas a otras.
 |
| Membranas y formación de capa hialina |
Un segundo efecto del aumento de Ca2+ es que provoca la activación del enzima NAD-Quinasa. Se encarga de transformar NAD en NADP. El óvulo va a necesitar sintetizar muchas cosas y el NADP es una molécula imprescindible para la biosíntesis.
El Ca2+ también activa una bomba de transporte que intercambia Na+ por H+. Este cambio desencadena las respuestas tardías. El cambio de pH es el que induce estas respuestas. Se forman proteínas, se activan sistemas de transporte, se divide el ADN. Entre 8 y 10 minutos después de la subida de pH en el interior, arrancan todos los procesos. Y una vez han arrancado, seguirán adelante y no se detendrán aunque el pH baje.
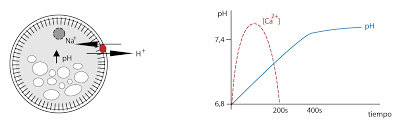 |
| Intercambios de calcio, sodio y cambios de pH en la fecundación. |
En resumen:
 |
| Resumen de la fase tardía de la fecundación. |
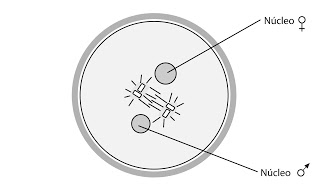
Cariogamia.
Si es una especie en la que el óvulo no ha terminado su maduración, esta debe completarse ahora, debe finalizar el proceso de meiosis. Cuando se completa, el proceso sigue.
El proceso pasa por varias fases. La primera fase es la monoáster, en la cual el núcleo masculino gira 180º, colocándose al revés, con el centriolo proximal hacia delante. Este se dividirá y comenzará a formarse el diplosoma. El núcleo masculino comienza a rehidratarse e hincharse, aumentando de tamaño y a descondensar la cromatina.
Posteriormente pasamos a la fase diaster. Los núcleos se van acercando y los diplosomas se han dividido y comienzan a separarse.
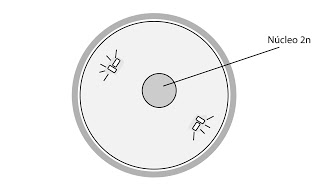
En la anfimixis o cariogamia propiamente dicha los núcleos se fusionan.
- Anatomía Celular: Retículo Endoplásmico
Se trata de uno de los sistemas de endomembrana más desarrollados. Dependiendo de la actividad de la célula, puede encontrarse con mayor o menor abundancia. Si bien no iene porque aparecier plenamente desarrollado en un momento determinado de la vida...
- Metabolismo: Cadena Respiratoria.
En entradas anteriores hemos analizado las fases iniciales del metabolismo de la glucosa: la glucolisis y la fosforilación oxidativa, junto con los mecanismos de degradación anaerobia y el Ciclo de Krebs. En este paso analizaremos la cadena respiratoria....
- Transporte A Través De Membrana
Transporte por difusión física simple. Es un transporte pasivo y no mediado. La membrana debe ser permeable para el producto en cuestión, que pasará desde la zona más concentrada a la zona más diluida. En teoría, cualquier molécula se puede disolver...
- Adn Mitocondrial: Significado Y Aplicaciones
Cuando ocurre alguna desgracia en la que toca hablar de identificación de cadáveres suele surgir un tema interesante y cuya importancia ha ido creciendo en los últimos años por diversas aplicaciones. Se trata de los análisis de ADN mitocondrial.¿Qué...
- Vih: El Virus Que Valió Un Nobel
Françoise Barre-SinoussiyLa academia sueca concedió en 2008 el premio Nobel de Medicina a Harald zur Hausen, Françoise Barré-Sinoussiy Luc Montaigner. Estos dos últimos son los investigadores que descubrieron el virus VIH (de las siglas en inglés,...