Biology
Aminoácido + ARNt + ATP ? Aminoacil-ARNt + AMP + 2Pi - ?G=0 Kcal/mol
La reacción está catolizada por la Aminoacil-ARNt sintetasa. La reacción tiene lugar en dos procesos, catalizados ambos por el mismo enzima.

- El Arn De Transferencia (arnt)
Los ARNt son los encargados de llevar los aminoácidos a su sitio de destino. Tendermos tantos ARNt como aminoácidos. Tanto en eucariotas como en procariotas se forman a partir de precursores más largos, con genes repetidos, unos a continuación...
- Arn Ribosomal Y Fabricación De Ribosomas
El ARN ribosomal (ARNr) constituye, junto con varias proteínas, los ribosomas. El ribosoma es la parte esencial de traducción del ARN. Los ribosomas de procariotas y eucariotas son ligeramente diferentes. Ambos están compuestos por dos subunidades,...
- Replicación De La Información Genética
Los estudios iniciales están hechos en células procariotas, sobre todo en Escheriricha coli. En la replicación intervienen veinte enzimas, aproximadamente, que forman un gran complejo denominado replisoma o sistema de replicasa. Y que iría replicando...
- Proteínas: Conformación Y Estructura.
Las proteínas son las piezas fundamentales de las células. Conforman el 50% del material intracelular. Tienen un gran abanico de funciones, intervienen en todos los procesos biológicos. Sus funciones fundamentales son: Catalizadores. Estructurales...
- Estructura Básica De Los Seres Vivos.
Aparentemente, todos somos capaces de diferenciar a un ser vivo de la materia inerte. Sin embargo, esto parte de error de asociar el concepto ser vivo al concepto de movilidad, crecimiento o cambio que no siempre se corresponde con la realidad. Una rápida...
Biology
Ribosomas: el proceso de traducción
Traducción.
Hasta ahora hemos visto el paso de ADN a ARN. El paso que estudiaremos ahora es el paso de ARN a polipéptidos, es decir, pasar de una secuencia de nucleótidos, una información lineal, a ua secuencia de aminoácidos que conformarán una información tridimensional.
Participan todos los tipos de ARN. El ARNm actuará como molde. El ARNt actuará como molécula adaptadora o decoficadora. El ARNr proporciona algunos centros catalíticos y ofrece el sitio o centro de enlace para que las unidades se coloquen bien.
El ARNm se leerá en la dirección 5??3?. La cadena de aminoácios crecerá del extremo -NH2 al extremo -COOH.
Y en muchas ocasiones se forman polisomas, varios ribosomas (ARNr) están leyendo a la vez un ARNm.
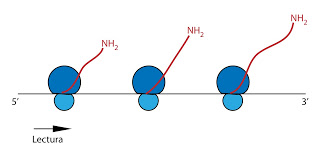 |
| Polisomas |
La lectura se lleva a cabo en una serie de etapas.
La primera etapa es la activación. Tiene lugar en el citoplasma. Los ARNt deben ir al citoplasma a través de los poros nucleares. Cada aminoácido se va aunir a un ARNt que le será específico. Algunos aminoácios poseen más de un ARNt.
Cuando el aminoácido se une al ARNt, el aminoácido se activa y se facilita la formaicón del enlace peptídico. Además, el ARNt es el adaptador, es el que correlaciona la secuencia, es la molécula traductora.
Usaremos el siguiente esquema para representar el ARNt con el aminoácido unido (es decir, al adaptador).
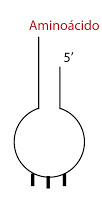 |
| ARNt con aminoácido |
El enlace específico es catalizado por las amioacil-ARNt-sintetasas. Existe al menos veinte diferentes, una por cada aminoácido, es decir, son específicas. Normalmente existen más de veinte, ya que como indicamos algunos aminoácidos tienen más de un ARNt.
La reacción global catalizada es:
La primera reacción es la siguiente:
Aminoácido + ATP ? Aminoácido-AMP + PPi
El aminoácido unido al AMP se denomina aminoacil-adenilato. Si vemos su fórmula química, sería como sigue:
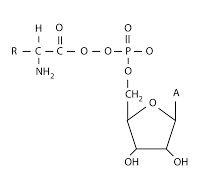 |
| Aminoacil-adenilato |
La segunda fase de la reacción sería la siguiente:
Aminoácido-AMP + ARNt ? Aminoácido-ARNt + AMP
Se trata de la transferencia del aminoácido al hidroxilo 3? del nucleótido terminal del ARNt. El paso se produce desde una posición 2 (en la que se encuentra al unirse al AMP) a un aposición 3 (en la que se encuentra cuando se une a la ribosa del ARNt).
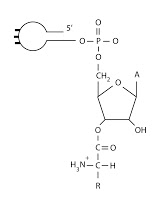 |
| ARNt con nucleótido unido |
Se forma un enlace con una ?G de 7Kcal/mol. Esta energía se recupera cuando la pirofosfatasa hidroliza el PPi, consiguiendo que la reacción se desplace hacia la derecha.
PPi + H2O ? 2 Pi - ?G=7 Kcal/mol.
Por cada aminoácido que se activa, se consume un ATP, pero esto equivale a dos enlaces fosfato, ya que también se consume la energía del enlace fosfato del PPi.
Ya tenemos unido el aminoácido al ARNt. El aminoácido solo va a ser identificado por el anticodon. Si se monta un aminoácido erróneo sobre el ARNt ya no se rconocerá el error. La corrección para evitar este problema se lleva a cabo en la primera de las dos reacciones. No obstate el enzima amonocetil-ARNt sintetasa es un enzima muy específico y se equivoca muy poco.
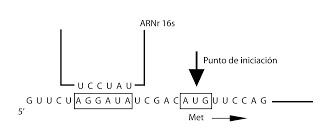
Ahora entrarímoas en la fase de iniciación. El ARNm tiene señales que le indican por donde tiene que empezar, tanto en los extremos 3? como 5? hay fragmentos que no se traducen.
Si son poligenómicos o policistrónicos, cada trozo codificante tendría sus señales de partida y sus señales de continuación. Las señales de inciación van a ser dos, una primera secuencia rica en bases púricas, que se empareja en el ribosoma con el ARNr 16s de la subunidad 20. Y una segunda que corresponde a una metionina, correspondiente a la señal de iniciación de la proteína. Esta secuencia de la metionina suele ser AUG, pudiendo sustituirse en algunos casos por la secuencia GUG.
En bacterias la metionina inicial no se corresponde a una metionina normal, sino a la fornilmetionina, F-Met. Por lo tanto, el primer ARNt será el correspondiente a este aminoácido especial, el F-Met-ARNtFMet. Se trata del ARNt iniciador. Solo se usa para comenzar las cadenas. Por lo tanto habrá dos ARNt para la metionina, el de la metionina normal, ARNtMet, y el que hemos indicado, para la F- Met iniciadora.
Estudiemos el inicio de la transcripción en porcariotas.
Para iniciar la transcripción, debe haber en el citoplasma una serie de factores protéicos disueltos: el IF-1, el IF-2 y el IF-3. El orden de actuación no está muy claro, aunque se supone que comenzarían entrando el IF-3 y el IF-1. La unión de estos dos separa las dos subunidades, o impide que se unan si ya están separadas. Ambos se unen a la subunidad 30s. Posteriormente entrarán el IF-2 unido a GTP, el ARNm y el F-Met-ARNtFMet. El IF-2-GTP se encarga de enlazar al F-Met-ARNtFMet y ponerlo correctamente. El IF-2-GTP solo reconoce al ARNt iniciador. Tras esta unión del ARNt y el IF-2-GTP se libera el IF-3, quedando la unidad 30s solo con el factor IF-1. A esta subunidad 30s con el ARNm, ARNt y demás factores unidos se le llama complejo de inciación 30s.
Ahora debe unirse la subunidad 50s. Al hacerlo, se liberan los factores IF-1 y IF-2 y el GTP se consume, generándose GDP+Pi. Es la subunidad 50s la que induce esta hidrólisis del GTP y la liberación de las IF-1 y IF-2. Tras el proceso, tenemos al ribosoma unido con sus dos subunidades, con el ARNm enlazado y el ARNt de inciación unido. Al conjunto se le denomina complejo de iniciación 70s.
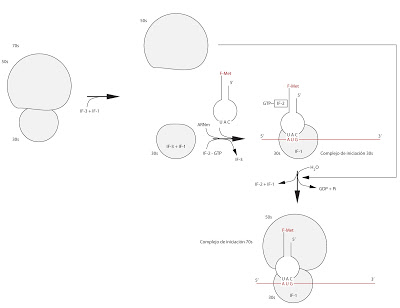 |
| Primera fase de la traducción |
El ribosoma tiene dos sutios de unión o locus, que es donde se posicionan los ARNt. Se denominan locus peptidilo o locus P y locus aminoacilo o locus A. Salvo en el caso de la adición del primer aminoácidos, que entra por el locus P, todos los demás entran al ribosoma por el locus A.
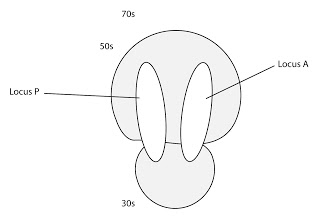 |
| Locus P y locus A del ribosoma |
Ahora comenzaría la fase de elongación. Partimos del complejo de iniciación 70s (donde lo dejamos en la iniciación). Para la elongación se requiere la adicción de nuevos EF, que se denominan EF-tu, EF-ts y EF-G.
Lo primero que sucede es que se coloca el siguiente aminoacil-ARNt en el locus A del ribosoma, que ahora se encuentra libre. Se coloca el ARNt que tenga el codon complementario a la cadena de ARNm y el correspondiente aminoácido unido a este ARNt. Este proceso tiene lugar con la colaboración del EF-tu. El EF-tu lleva unido una molécula de GTP y reconoce a cualquier ARNt excepto al iniciador. Posteriormente actuará el enzima Peptidil transferasa, presente en la subunidad 50s, formándose el enlace peptídico. La reacción tiene lugar entre el grupo carboxilo del aminoácido esterificado (inicial) que se encuentra en el locus P con el grupo amino del aminoácido que se encuentra en el locus A.
Durante la reacción el se consume el GTP que se encuentra unido al EF-tu, transformándose en GDP y obteniéndose por lo tanto EF-tu-GDP. Este GTP debe ser repuesto. De esto se encarga el EF-ts, que colabora en que otro GTP se transforme en GDP y de esta forma el EF-tu-GDP pase a ser de nuevo EF-tu-GTP.
Tras la reacción de esterificación, los dos aminoácidos se quedan enganchados al ARNt del segundo aminoácido, en el locus A.
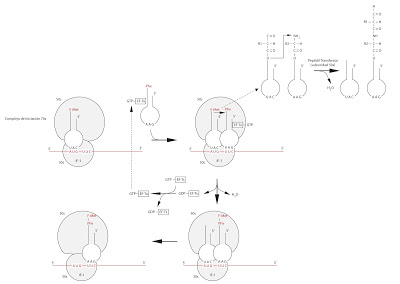 |
| Segunda fase de la traducción |
El siguiente paso es la traslocación. El ARNt que poseía la metionina inicial y que ahora no tiene aminácido se desprende, dejando el locus P vacío. Y ahora el peptidil-ARNt, es decir, el ARNt que lleva unidos los dos aminoácidos, debe pasar del locus A al locus P. Lo hace gracias al EF-G. Este lleva unido GTP, es decir, es un EF-G-GTP y durante el proceso se consume este GTP, obteniéndose GDP, PPi y liberándose el EF-G.
Tras la traslocación tenemos al peptidil-ARNt en el locus P y el locus A vacío. Se trata de una situación equivalente a la de partida. A partir de ahora, tantos aminoácidos como tenga la cadena, tantas veces como se deben repetir todos estos pasos.
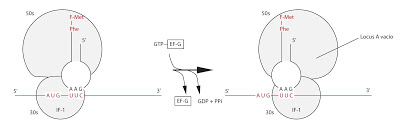 |
| Traslocación |
Y llegamos a la terminación. Tras todo el proceso, al final de la secuencia que codifica para la proteína, vendrá una de las tres señales que codifican para la señal de parada o señal de stop. Normalmente no existe un ARN con anticodón complementaria para esta señal. Como no hay ARNt, se para. Pero debe soltarse el polipéptido. Se necesita algún factor de terminación, que se denominarán RF (en Escherichia coli hay tres factores de terminación). Estos factores de terminación indicuen que se rompa en elnace que une al polipéptido con el ARNt al que llevan unidos desde el inicio de la traducción. Y tras el proceso, se liberarán el ARNt libre del polipéptido, el polipéptido y el ARNm.
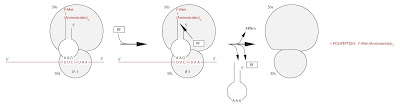 |
| Terminación |
Ahora el polipéptido debe tomar una conformación. Está esencialmente dirigida por su secuencia y el medio en el que se traduce. Es frecuente que esta conformación se vaya adquiriendo a la vez que se va traduciendo, de acuerdo con el medio en que se encuentra.
Modificaciones post-traduccionales.
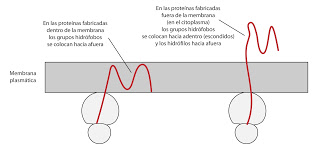
Cuando los ribosomas se encuentran adosadas a las membranas, se enganchan por la subunidad grande.
En ocasiones, el polipéptido necesita modificaciones post-traduccionales para que sea funcional. En bacterias, el grupo fornilo se elimina y en ocasiones junto con este grupo fornilo se eliminan varios aminoácidos más.
Debemos tener en cuenta, además, que algunas proteínas poseen aminoácidos que no tienen traducción y deben añadirse o modificarse después de la traducción. Deben añadirse grupos. Y completar otros aspectos, como por ejemplo los enlaces disulfuro entre los radicales.
- El Arn De Transferencia (arnt)
Los ARNt son los encargados de llevar los aminoácidos a su sitio de destino. Tendermos tantos ARNt como aminoácidos. Tanto en eucariotas como en procariotas se forman a partir de precursores más largos, con genes repetidos, unos a continuación...
- Arn Ribosomal Y Fabricación De Ribosomas
El ARN ribosomal (ARNr) constituye, junto con varias proteínas, los ribosomas. El ribosoma es la parte esencial de traducción del ARN. Los ribosomas de procariotas y eucariotas son ligeramente diferentes. Ambos están compuestos por dos subunidades,...
- Replicación De La Información Genética
Los estudios iniciales están hechos en células procariotas, sobre todo en Escheriricha coli. En la replicación intervienen veinte enzimas, aproximadamente, que forman un gran complejo denominado replisoma o sistema de replicasa. Y que iría replicando...
- Proteínas: Conformación Y Estructura.
Las proteínas son las piezas fundamentales de las células. Conforman el 50% del material intracelular. Tienen un gran abanico de funciones, intervienen en todos los procesos biológicos. Sus funciones fundamentales son: Catalizadores. Estructurales...
- Estructura Básica De Los Seres Vivos.
Aparentemente, todos somos capaces de diferenciar a un ser vivo de la materia inerte. Sin embargo, esto parte de error de asociar el concepto ser vivo al concepto de movilidad, crecimiento o cambio que no siempre se corresponde con la realidad. Una rápida...