Biology
Alton Brown from Food Network hates a unitasker. He wants all his kitchen tools to have more than one function ? I least I think it?s just his kitchen tools. But he might just as well be talking about biology. Nature hates a unitasker, that?s why some many things in our cells have multiple jobs.
The king of the moonlighting proteins is glyceraldehyde -3-phosphate dehydrogenase (GAPDH). Sure, it's one of the enzymes that breaks glucose down to pyruvate, but it does so much more ? like helping to maintain the ends of our chromosomes (telomeres), working to move tRNAs out of the nucleus, controlling the expression of some genes, especially those involved with gamma-interferon, repairing our DNA when it is damaged. And apparently GAPDH is crucial for helping cells to bring in particles from outside (endocytosis). That?s a full day.
The basal body is about 100 nm wide and 150 nm long, and serves as the base of the flagellum or cilium. If a cell has hundreds of cilia, like the male gametes of the cycads we talked about last week, then it has hundreds of basal bodies as well ? one per cilium.
If you remove the centrioles after S phase, most cells can still go through mitosis just fine. In fact, there many cell types that don?t have centrioles at all (higher plants, some protists, most fungi). Even in cells that are supposed to have centrosomes, destroying centrioles with a laser after S phase doesn?t always affect mitosis negatively.
After centriole duplication by prescribed pathways, the basal body matures. A 2011 review shows that there are many proteins involved in the process. First the distal end of the centriole is capped, then it migrates to the cell membrane and docks. A transition zone is produced that allows for selective movement of molecules up and down the inside of the axoneme (intraflagellar transport, IFT).
The basal body does a different job by controling the direction of movement of the cilium or flagellum! The prokaryotic flagellar motor spins the prokaryotic flagella as we have described, but eukaryotic cilia or flagella beat, rather than flop around, and the direction in which they beat is important. Cilia on a cell beat in concert in one direction, and the basal body is asymmetrical enough to drive this directional beating. Believe it or not, this is controlled by a protein called disheveled.
Cells without either centrioles and basal bodies suggest that basal body function came first; since you can?t have undulipodia without basal bodies, but our discussion above shows you can have mitosis without centrioles.
Henderson, B., & Martin, A. (2014). Protein moonlighting: a new factor in biology and medicine Biochemical Society Transactions, 42 (6), 1671-1678 DOI: 10.1042/BST20140273
Kobayashi, T., & Dynlacht, B. (2011). Regulating the transition from centriole to basal body The Journal of Cell Biology, 193 (3), 435-444 DOI: 10.1083/jcb.201101005
Debec, A., Sullivan, W., & Bettencourt-Dias, M. (2010). Centrioles: active players or passengers during mitosis? Cellular and Molecular Life Sciences, 67 (13), 2173-2194 DOI: 10.1007/s00018-010-0323-9
Boisvieux-Ulrich E, & Sandoz D (1991). Determination of ciliary polarity precedes differentiation in the epithelial cells of quail oviduct. Biology of the cell / under the auspices of the European Cell Biology Organization, 72 (1-2), 3-14 PMID: 1756309
- Animal Eukaryotic Cell Components: Structure And Function
I posted a picture of the eukaryotic animal cell earlier, but it was simply a labeled diagram. Here we will go into each of the major parts of the cell, as well as each organelle's function, not to mention the overall structure of the cell. As the...
- Mitosis
Prophase Chromatin condenses to form chromosomes. Centrioles migrate to opposite poles of the cells (except in plant cells which do not have centrioles). Microtubules develop and form a star shaped structure known as an aster, and spindle fibres may develop....
- #32 Summary Of Cell And Nuclear Division
1. Growth of a multicellular organism is a result of parent cells dividing to produce genetically identical daughter cells. 2. During cell division the nucleus divides first, followed by division of the whole cell. 3. Division of a nucleus to produce...
- Cell Organelles In Eukaryotes
Name of the cell organelle Functions 1. Cell wall (Absent in animals and some protists. Present in bacteria, fungi, algae and plants). - Gives rigidity and support to the tissues and organs. Gives characteristic...
- A Tale Of Two Tails
Biology concepts ? flagella, bacteria, prokaryotes, eukaryotes, undulipodia, axoneme, basal body, centriole Everyone has the dream where you show up for a class that you didn?t know was on your schedule, only to be having a test. But in second place...
Biology
Evolving A Second Job
Biology concepts ? protein moonlighting, undulipodia, evolution, basal body, centriole, GAPDH, intraflagellar transport
 |
| Today?s post is on a multitasking cell structure. This would make Alton Brown proud, since he hates tools that do only one thing. The University of Miami of Florida football team runs through fire extinguisher blasts when they enter the stadium ? maybe Alton can find a second use for his. |
This phenomenon is called protein moonlighting. The re-evaluation of the human genome (about 19,000 genes) suggests that many proteins have more than one distinct function. This would allow for a relatively small number of genes to provide a large functional proteome (the total number of protein functions). As such, a 2014 study is showing the importance of moonlighting proteins in health and biology.
There are rules for a protein to have a legitimate second job. It?s only moonlighting if the two functions are unrelated, the functions can't be carried out by two different domains of the protein either. This would suggest a gene fusion event. The two functions must be independent, so ablating one doesn?t affect the other.
There are hundreds of examples of moonlighting proteins in the literature now, and more are sure to follow. There are examples aplenty within the glycolysis pathway; you know, the breakdown of sugar for energy. No fewer than seven of the ten glycolytic enzymes are known to have other jobs.
 |
| Crystallin proteins (alpha and beta) make up the majority of the lens and cornea of the eye. They are transparent, but they do more than that. Recent studies show that they have enzymatic activity in other tissues. Aldehyde dehydrogenase and transketolase are enzymes that turn out to be moonlight crystallins. |
But as amazing as GAPDH is, today?s example of a multitasker is even more rare, in that the moonlighter is a complete structure, made of many proteins, and has two distinctly different jobs. What?s more, each function has its own exceptions. What's our structure of interest? The basal body, or perhaps I should call it the centriole.
Let?s talk about the basal body first. This is the base of the eukaryotic undulipodia (cilia and flagella). These moving tails come in two parts; the basal body and the axoneme. We talked at length about the axoneme a few months ago, with its nine doublet microtubules surrounding two singlet microtubules (9[2]+2, see picture below). Undulipodia movement, as opposed to the motor driven prokaryotic flagella, is achieved by sliding the different doublets forward and back past one another.
But in those previous posts we didn?t talk much about the basal body. It too is a ring of microtubules, although these are shorter polymers that in the axoneme. Instead of doublets, there are almost always nine sets of triplet microtubules, and there are no center microtubules (9[3]+0). Of course, there were a couple of exceptions, and we talked about them.
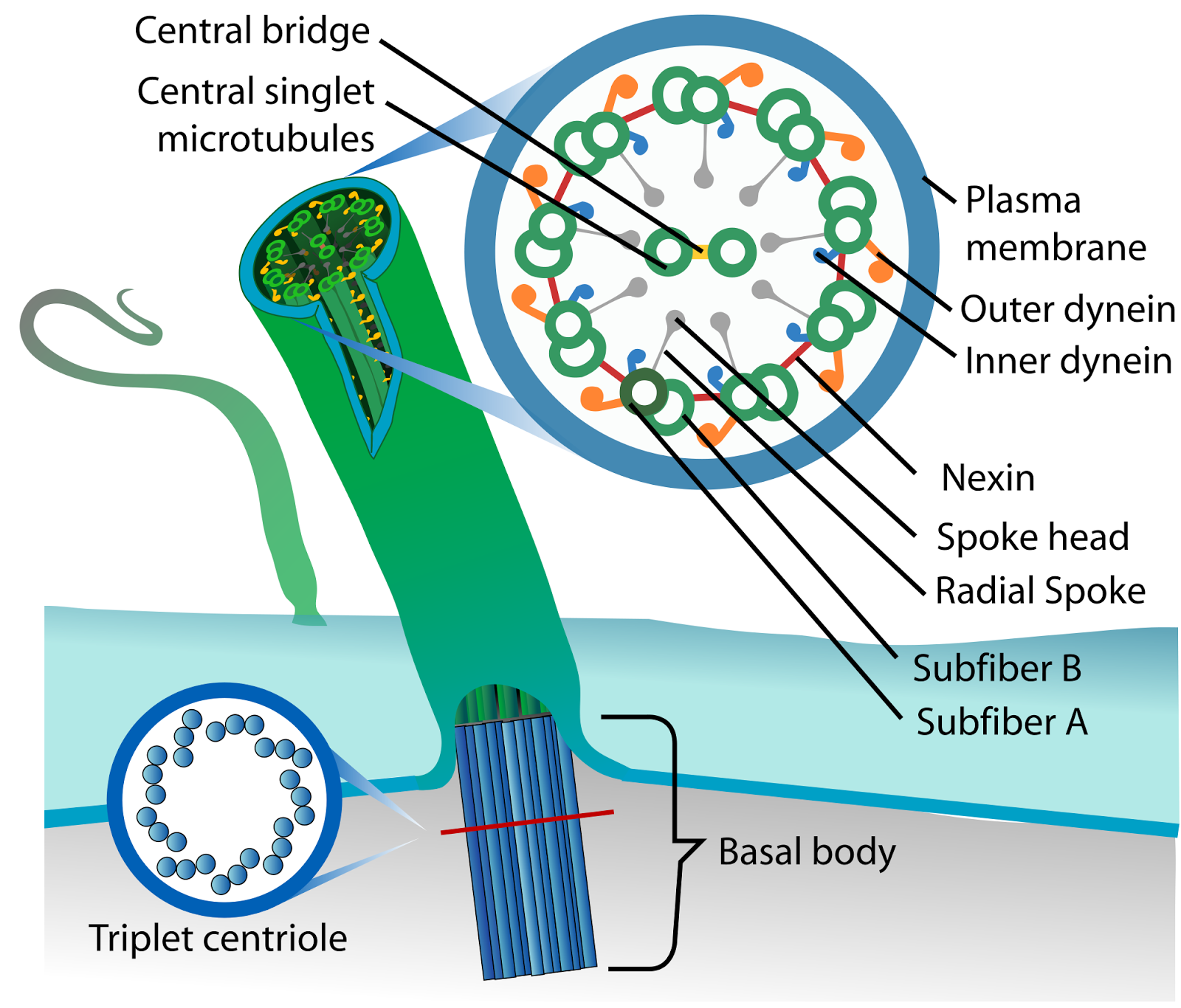 |
| The basal body has a 9(3) + 0 structure, while the axoneme is 9(2) + 0. While this cartoon shows the complexity of the axoneme, our post today highlights the complexity of the basal body. One thing this cartoon does show, the axoneme is sheathed in the plasma membrane, it isn?t a protein structure sticking out through the membrane. |
The basal body serves as the nucleation site for mictrotubule growth into the axoneme. It?s like a skyscraper, the microtubule girders are built vertically on the basal body foundation; only here, the basal body sparks a self-assembly of the microtubules. You don?t need fearless guys climbing the beams to build an axoneme.
If we turn our attention to the centriole, we find that it?s used in mitosis. When a cell divides, each progeny cell receives one centrosome. Don?t confuse the centrosome with a centriole or a centromere (a near center point of a chromosome, it holds the two chromatids together). The centrosome is more of an area, it contains two centrioles, a mother and a daughter, and the pericentriolar matrix (PCM) amorphous group of proteins that help the centrioles do their job.
Each centriole is a complex microtubule formation in a 9(3)+0 arrangement, about 120 nm wide and 175 nm long. This is exactly the structure of the basal body ? they?re the same thing! Well, almost.... a centriole has to mature into a basal body.
During S phase of the cell cycle (when the chromosome are replicated) the centrosome will duplicate. Each centriole grows another one from its side, at a right angle. The mother is a mother again, and the daughter becomes a mother for the first time. The two mother/daughter pairs then gather their own PCMs and move to the sides of the nucleus. When mitosis time comes, microtubules grow toward the chromsosomes, but from the mother centriole only. This is the spindle and will help pull the chromatids apart during cell division.
 |
| Start at the top left. During the cycle, a pair of centrioles will separate and each will grow another from the proximal end. The daughter centrioles then have to mature, with proteins added toward the distal end. During mitosis, they two pairs separate and form the spindle body, which pulls the chromatids apart. |
A commentary published in 2010 talks about how many cell types, including some oocytes can undergo meiosis without centrioles, while centriole numbers than are re-established after fertilization. However, in other tissues, loss of centrioles leads to genetic instability over time, even if the spindle will develop without the centrosome. To this point, we still haven?t resolved the issue of whether centrioles are necessary for proper cell division.
In the vast majority of cases, centrioles come from centrioles. One serves as a template to form the second. The process through which this occurs has just begun to be revealed in the past few years. There is a linking fiber from the proximal end of the mother centriole that acts as a seed point to start aggregation of the daughter centriole at a 90? angle to the first.
The process is very complicated, but is carried out spontaneously, without specific gene products to guide it. It was first believed that if centrioles were lost, then they could not be regained. Of course, they also thought that centrioles had their own DNA. Now we know that in cases where centrioles are lost, they can form de novo, and function just fine in mitosis or as basal bodies. In several studies, removal of centrioles or cells without centrioles to begin with allows for new centrioles being formed from aggregated microtubules.
 |
| Intraflagellar transport is the method by which the axoneme grows, adding tubulin monomers to the end. The proteins are carried up the axoneme by walking proteins. These are important not for just cilia assembly, but also for cell signaling and sensing that occurs in the undulipodia. |
Finally, there is attachment of accessory bodies like rootlets to anchor it to the membrane, transition fibers to move to the axoneme and distal appendages. Only after all this maturation of the basal body can the axoneme be built by IFT. This is how the process happens in all species EXCEPT fruit fly male gametes and the microbe Plasmodium yoelii, where the axoneme grows BEFORE plasma membrane docking of the basal body.
What?s more, you can always reverse it and go from basal body back to centriole, so their functions must somewhat overlap. The basal body and centriole both serve as microtubule organizing centers (MTOCs). So what makes it a moonlighting structure?
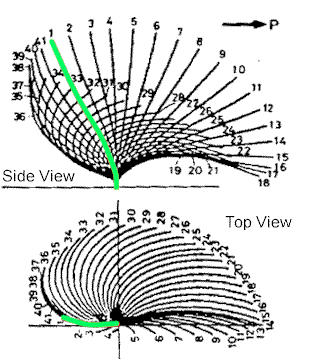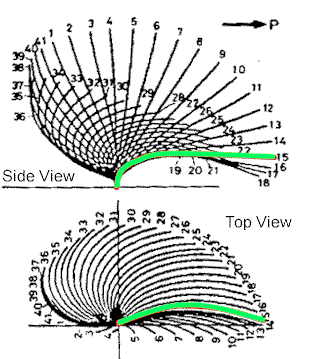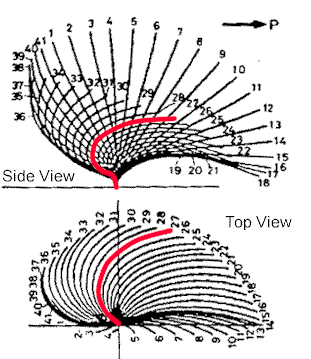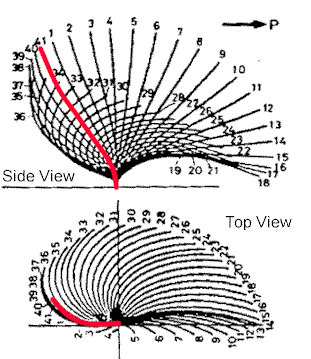 |
| Prokaryotic flagella just whip around in a circle, but eukaryotic undulipodia have more of a beating motion. The side view shows how the flow is one direction, while the top view indicates that the motion is circular, but not like a propeller. |
That?s a little weird, but what?s really weird is the kids. In a 1991 paper, part of the oviduct of a quail was reversed, so that the cilia beat toward the ovary instead of toward the uterus. When cells divided through embryo and chick development, the cilia of the progeny cells had the same orientation as those in the parent! They continued to beat in the wrong direction. Since basal bodies are duplicated from basal bodies, the mother basal bodies gave rise to daughters that beat the same direction.
The evidence presented shows that we have an ancient, yet complex, structure that has a couple of important jobs. The question for evolutionary biologists is which came first, the basal body or the centriole? The last common eukaryotic ancestor (LECA), the cell from which all eukaryotic cells descend, had undulipodia, so the basal body is an extremely old structure. But all eukaryotic cells undergo mitosis or meiosis, so the centriole must be important too.
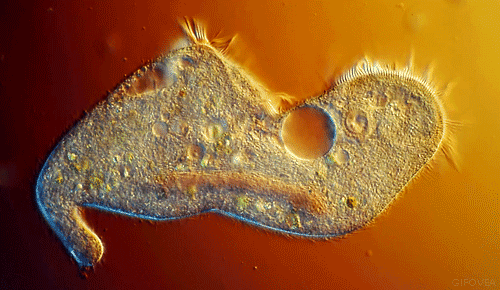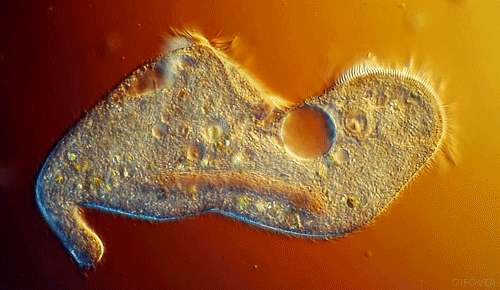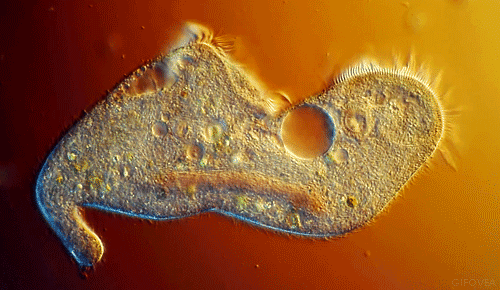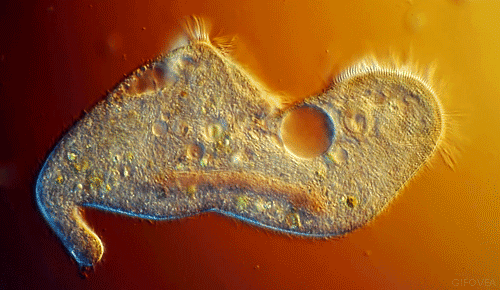 |
| Notice the cilia that are the upper most in the animation. They show best the coordinated beating that pushes fluid in one direction. |
Cells without either centrioles and basal bodies suggest that basal body function came first; since you can?t have undulipodia without basal bodies, but our discussion above shows you can have mitosis without centrioles.
On the other hand, centrioles have to mature to become basal bodies, wouldn?t this suggest that they came first? Or perhaps the basal body was the primary product and nature managed to find a use for one if its precursors. What do you think, is the basal body the chicken or the egg?
Next week let?s look at one of the animal kingdoms great exceptions in terms of cilia, even though primitive and higher animals have them, round worms don?t cilia or flagella!
For more information or classroom activities, see:
Protein moonlighting ?
http://www.slideshare.net/vimalacbr/protein-moonlighting
http://www.biochemistry.org/TabId/379/MeetingNo/SA162/view/Conference
http://www.ebi.ac.uk/interpro/potm/2007_5/Page2.htm
http://www.moonlightingproteins.org/
http://www.cvent.com/events/the-biological-and-biomedical-consequences-of-protein-moonlighting/event-summary-31ba211f647f4cdda499019b008c64da.aspx
http://www.biosyn.com/tew/protein-moonlighting.aspx
http://www.petercollingridge.co.uk/blog/proteins-look-enzymes-arent
Centriole ?
http://www.cellsalive.com/cells/cell_model.htm
http://www.scienceteacherprogram.org/biology/savitzky06.html
http://vimeo.com/58347006
http://www.discovery.org/multimedia/video/2010/02/centriole-animation/
https://www.youtube.com/watch?v=L9DbrOm3zj0
https://www.youtube.com/watch?v=ATlUv-AGhEU
http://www.biology4kids.com/files/cell_centriole.html
http://bscb.org/learning-resources/softcell-e-learning/centriole/
http://micro.magnet.fsu.edu/cells/centrioles/centrioles.html
https://sites.google.com/a/asu.edu/the-almighty-cell/the-source/animal-cell/centriole
http://www.basic.northwestern.edu/g-buehler/eyes.htm
http://www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb2/part1/microtub.htm
Basal body ?
http://classroom.synonym.com/basal-bodies-form-cilia-flagella-originate-from-23403.html
http://www.ncbi.nlm.nih.gov/pubmed/19147000
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/C/Cilia.html
http://scienceblogs.com/transcript/2007/05/22/a-note-on-centrioles-basal-bod/
Centrosome ?
http://classroom.synonym.com/difference-between-centriole-centrosome-13002.html
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/C/Centrioles.html
https://www.princeton.edu/~achaney/tmve/wiki100k/docs/Centrosome.html
http://events.embo.org/14-centrosome-spb/
http://www.biomedcentral.com/1741-7007/11/28
https://www.youtube.com/watch?v=eRfTQ1Pptys
http://bcs.whfreeman.com/lodish5e/content/cat_010/01010-03.htm?v=chapter
https://prezi.com/ey2ffxepphzl/centrioles-and-centrosomes/
http://www.sciencephoto.com/media/526790/view
- Animal Eukaryotic Cell Components: Structure And Function
I posted a picture of the eukaryotic animal cell earlier, but it was simply a labeled diagram. Here we will go into each of the major parts of the cell, as well as each organelle's function, not to mention the overall structure of the cell. As the...
- Mitosis
Prophase Chromatin condenses to form chromosomes. Centrioles migrate to opposite poles of the cells (except in plant cells which do not have centrioles). Microtubules develop and form a star shaped structure known as an aster, and spindle fibres may develop....
- #32 Summary Of Cell And Nuclear Division
1. Growth of a multicellular organism is a result of parent cells dividing to produce genetically identical daughter cells. 2. During cell division the nucleus divides first, followed by division of the whole cell. 3. Division of a nucleus to produce...
- Cell Organelles In Eukaryotes
Name of the cell organelle Functions 1. Cell wall (Absent in animals and some protists. Present in bacteria, fungi, algae and plants). - Gives rigidity and support to the tissues and organs. Gives characteristic...
- A Tale Of Two Tails
Biology concepts ? flagella, bacteria, prokaryotes, eukaryotes, undulipodia, axoneme, basal body, centriole Everyone has the dream where you show up for a class that you didn?t know was on your schedule, only to be having a test. But in second place...